Introduction
Ischemic cerebrovascular disease is a common clinical disease with high disability rate, high recurrence and slow recovery [1]. The mortality rate of ischemic cerebrovascular disease is second only to heart disease and is one of the major causes of death in humans [2, 3]. Focal cerebral ischemia-reperfusion injury is a complex pathological cascade. The factors affecting this pathological process are various, including the release of inflammatory mediators after ischemia, endothelial cell dysfunction, mitochondrial damage and oxygen failure [4, 5].
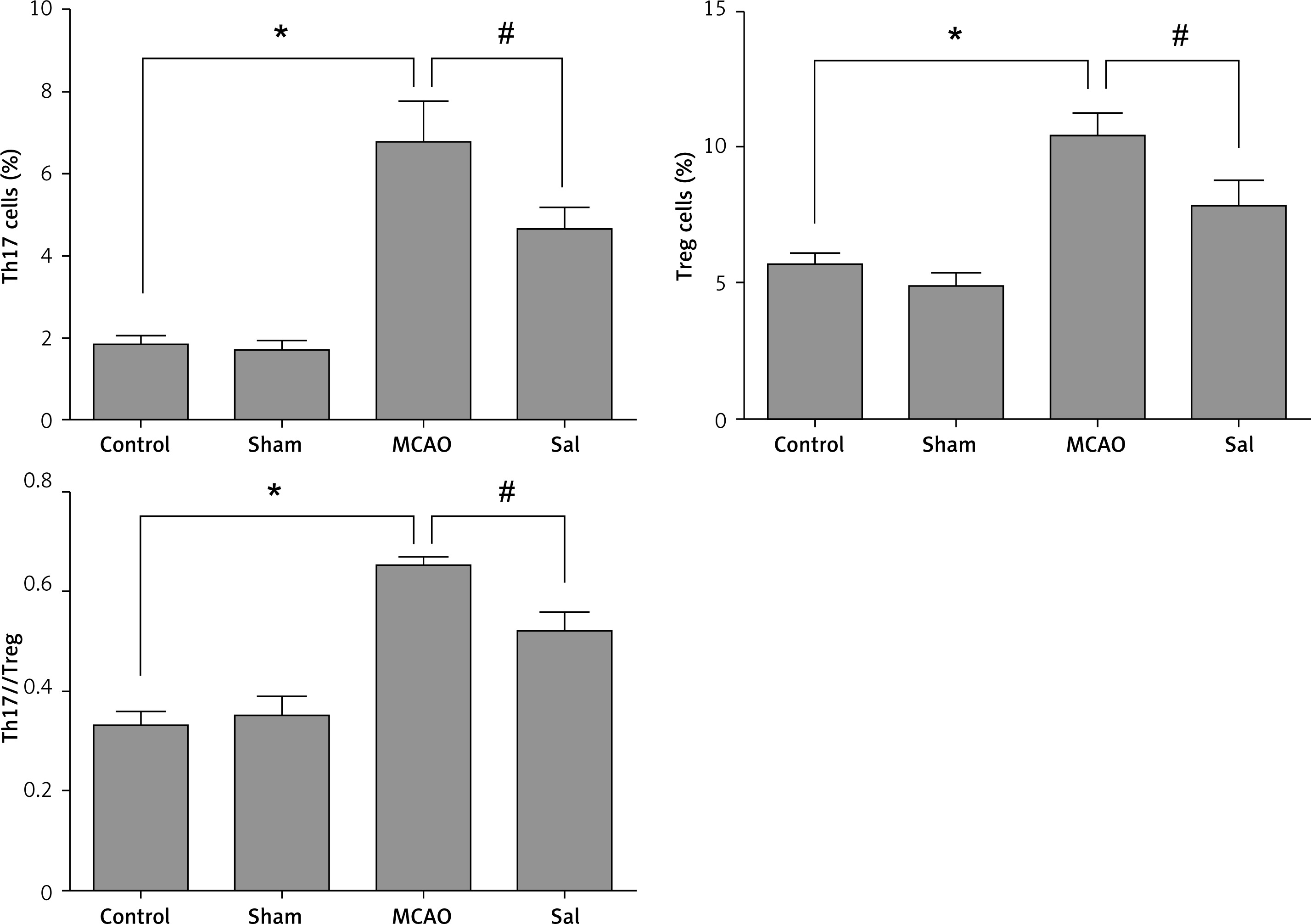
Th17 cells are CD4+ T cells that secrete interleukin (IL) 17 specifically under the regulation of the nuclear transcription factor ROR-γt. IL-17 can promote the differentiation and maturation of neutrophils by inducing granulocyte colony-stimulating factor, stimulate the activation of monocytes and fibroblasts, and exert a potent pro-inflammatory effect [6, 7]. Therefore, IL-17 is considered to be a key factor in inducing an inflammatory response and is involved in a variety of autoimmune diseases [8]. Th17 cells not only secrete IL-17, but also secrete cytokines such as IL-21 and IL-22, and participate in various autoimmune diseases. Treg cells are a subpopulation of CD4+ T cells with immunosuppressive activity [9]. Treg cells inhibit the production of inflammatory cytokines by releasing the cytokines IL-10 and TGF-β to inhibit the function of T cells and antigen presenting cells [10]. Foxp3 is an important transcription factor of Treg, and its sustained expression is a key factor in maintaining Treg inhibitory activity [11]. Treg exerts immunomodulatory effects by up-regulating the expression of inhibitory immune cell surface molecules and down-regulating the expression of genes involved in activated T cells [12]. Th17/Treg balance is important for focal cerebral ischemia/reperfusion injury. Once the Th17/Treg balance is destroyed, it will cause disease. Therefore, maintaining Th17/Treg balance is important for the treatment of focal cerebral ischemia/reperfusion injury. Studies have shown that IL-17 can promote the secretion of IL-6, TNF-α and other pro-inflammatory factors, by antigen-presenting cells, and participate in and expand the host inflammatory response [13–15]. IL-6 is a key cytokine that can determine the differentiation of naive T cells into Th17 or Treg cells. The process involved in the induction of Th17 cell differentiation may be closely related to the activation of the transcription factor STAT-3 [16].
Rhodiola is the dry rhizome of Rhodiola rosea L., and salidroside (Sal) is the main active ingredient of Rhodiola [17]. In the rat model of focal cerebral ischemia/reperfusion injury, salidroside can reduce the infarct volume of the rat brain and reduce the neurological deficit score [18], but the exact mechanism is still unclear. The aim of this study was to investigate the effects of salidroside on the balance of Th17/Treg cells in rat brain tissue induced by focal cerebral ischemia-reperfusion injury, as well as the expression levels of IL-6, TNF-α, MCP-1, STAT-3 and NFκ-B2 protein, and to further explore the mechanism of action of salidroside on neuroprotection in middle cerebral artery occlusion (MCAO) model rats.
Material and methods
Experimental grouping and establishment of MCAO model rats
The rats were purchased from the experimental animal centre of Southern Medical University. Forty rats were randomly divided into 4 groups – control group, sham group, MCAO group, Sal group – with 10 rats in each group, and kept in cages. The establishment of MCAO model rats followed the following steps: rats were anesthetized by intraperitoneal injection of 10% chloral hydrate (3 ml/kg body mass), methylpolysiloxane-coated nylon monofilaments were inserted from the left common carotid artery to the beginning of the middle cerebral artery. Two hours after embolization, the nylon monofilament was pulled out. The middle cerebral artery was not embolized in sham-operated animals. Before the operation, the rats were fed for 10 days to adapt to the environment, and the light was 12 h cycles during the day/night. At 3 days before the operation, the Sal group rats were given an intraperitoneal injection of Sal (Meilunbio, China) 12 mg/kg once a day, while the rest of the group was given an intraperitoneal injection of equal isotonic saline. During the operation, the temperature of the animal was maintained at 37°C. On the second days after surgery, the peripheral blood mononuclear cells were isolated and the count of Th17 and Treg cells was detected by flow cytometry.
Evaluation of neurologic deficit and detection of infarct size
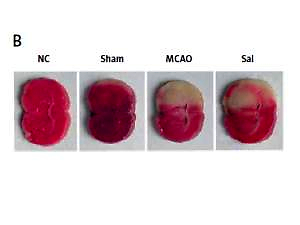
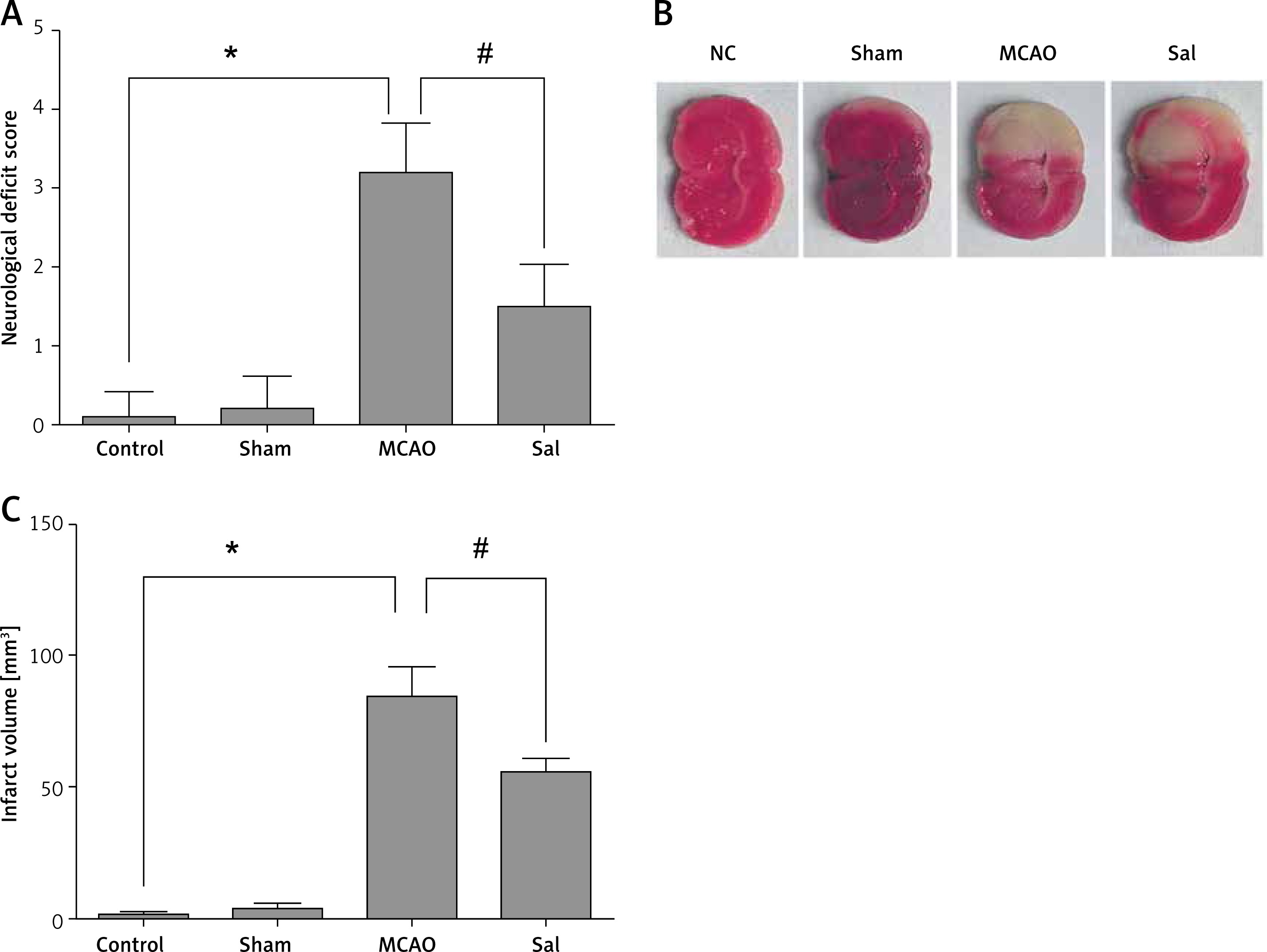
According to the Longa assessment [19], on the second day after surgery, the degree of neurologic deficit was evaluated in rats: grade 0, no defect; grade 1, no extension of the lateral forelimb; grade 2, lateral forelimb flexion; grade 3, mild reverse circle; grade 4, serious circle; grade 5, contralateral paralysis. Rats were killed and brain tissues were collected. Some of them were frozen in liquid nitrogen and transferred to –80°C. The other part was cut into 3 mm by slicers and stained with 1% TTC for 30 min. After staining, the film was scanned and photographed, and the infarct size was analyzed by ImageJ software.
Apoptosis in rats’ brain cells
Rat brain tissue was cut into 4 mm slices and stained according to the TUNEL kit instructions provided by reagents (Beyotime, China). Five visual fields were randomly selected to count positive staining cells under a 100× microscope. The apoptosis was expressed by the ratio of TUNEL staining positive cells to the total number of cells in the field of view.
Cell culture and siRNA interference
Rat spleen cells of MCAO model rats were separated according to a method described in the literature [20]. The cytokine TGF-β (2 ng/ml) was added to induce Treg cells, and cytokines IL-6 (20 ng/ml) and TGF-β (2 ng/ml) were added to induce Th17 cells according to the reference materials [21]. The cells were divided into four groups: control group (control), the hypoxia group (H), the hypoxia + Sal group (HS), the siRNA + hypoxia + Sal group (SHS). The cells were cultured at 37°C, 5% CO2 culture box. After the state became stable, the cells were used for gene silencing experimentation. Before receiving the drug treatment, the cells were treated with SATA-3 siRNA in accordance with the instructions provided by the manufacturer (Guangzhou Ribobio). The cells used in the hypoxia experiment were incubated with Sal (10–4 mol/l) for 24 h under the conditions of 1% O2, 5% CO2, and 94% N2. The treated cells were divided into 3 parts: the first part was used to detect cell apoptosis, the second part of the cells was used to detect cell SOD and MDA, and the third part was used to detect the protein expression.
MTT experiment
The cell density was adjusted to 1 × 105/ml and inoculated with 100 μl per hole in a 96-hole plate. After the cell growth reached 70% confluence, the cells were grouped by the above method. After the reoxygenation was finished, the cells were incubated with 20 μl of MTT (Beyotime, China) for 4 h. After centrifugation the supernatant was discarded, then 150 μl DMSO was added to each hole and shaken for 10 min. After the crystals were dissolved, the OD values of each group were detected at the wavelength 490 nm.
Flow cytometry
For the expression of Th17 and Treg cells, peripheral blood mononuclear cells were isolated and the count of Th17 and Treg cells was detected by flow cytometry according to the reference [22]. For the apoptotic, after recovery, the cells were washed with PBS buffer 3 times. The cells were stained with FITC and PI for 10 min at room temperature. The apoptotic cells were detected by flow cytometry and the percentage of apoptotic cells was counted.
Detection of cell SOD and MDA
Changes of SOD activity and MDA content can sometimes occur due to acute stress and there is increased enzyme activity to eliminate free radicals that have arisen. After recovering the treated cells, the MDA in the medium was detected by the Lipid Peroxidation MDA Assay kit and SOD in the medium was detected with the Total Superoxide Dismutase Assay kit based on the instructions provided by the manufacturer. SOD activity and MDA content were detected with the method as reported previously [23]. All tests were repeated 3 times, and the average value of the test data was obtained.
Western blot
50 mg tissue samples or 106 cells were taken from each sample. The total protein was extracted according to the instructions provided by reagents (Beyotime, China). A 60 μg protein sample was taken for SDS-PAGE electrophoresis. The protein was transferred to the polyvinylidene fluoride (PVDF) membrane. The corresponding antibody was added to the TBST closed buffer to incubate overnight at 4°C. The second antibody was added and incubated for 1 h at 37°C. The results were observed after exposure to chemiluminescence. GAPDH was used as the internal reference, and Bio-image (Bio-Rad) analysis software was used to collect signals and grayscale scanning.
Statistical analysis
Statistical analysis was performed using SPSS 19.0 (IBM Corp., Armonk, NY, USA) and GraphPad PRISM version 5.00 (GraphPad Software, Inc., La Jolla, CA, USA) software. Data are presented as mean ± standard deviation (SD), determined using single factor analysis of variance (ANOVA). Statistical significance was evaluated by one-way ANOVA followed by the least significant difference test or Dunnett’s T3 post hoc analysis. P < 0.05 was considered to indicate a statistically significant difference.
Results
Sal can reduce neurological deficit and infarct size in MCAO rats
As shown in Figure 1 A, compared with the sham group, the MCAO group showed poor neurological function, while Sal could significantly reduce neurological deficit. The ischemic lesion area was not detected in the sham group, while in the MCAO group, the infarct size was significantly increased. However, after Sal pretreatment, infarct size decreased significantly (Figures 1 B, C). These results indicate that Sal can reduce infarct size induced by ischemia in rats.
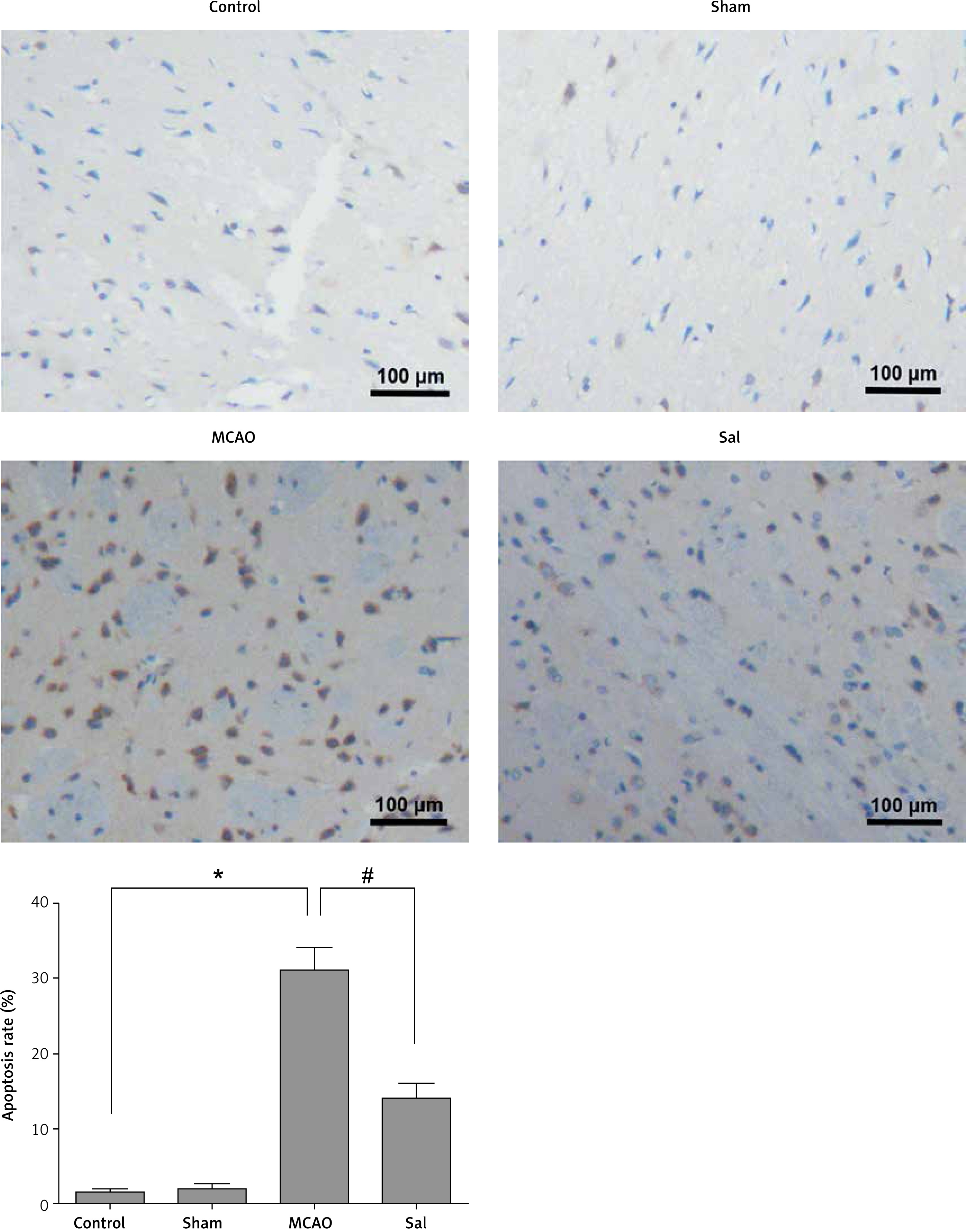
Sal can inhibit apoptosis of brain cells in MCAO rats
As shown in Figure 2, the apoptosis of brain cells in the MCAO group was significantly higher than that in the control group. However, the Sal treatment can significantly reduce the apoptosis of brain cells. These results indicate that Sal can inhibit the apoptosis of brain cells in MCAO rats.
Figure 2
Apoptosis was detected with the TUNEL assay in rats’ brain cells. Compared with control groups, MCAO group rats showed a significant increase in apoptosis, but the opposite results appeared in Sal groups. Dark brown represents apoptotic cells and the blue denotes nuclei
Compared with the control group, *p < 0.05; compared with MCAO group, #p < 0.05.
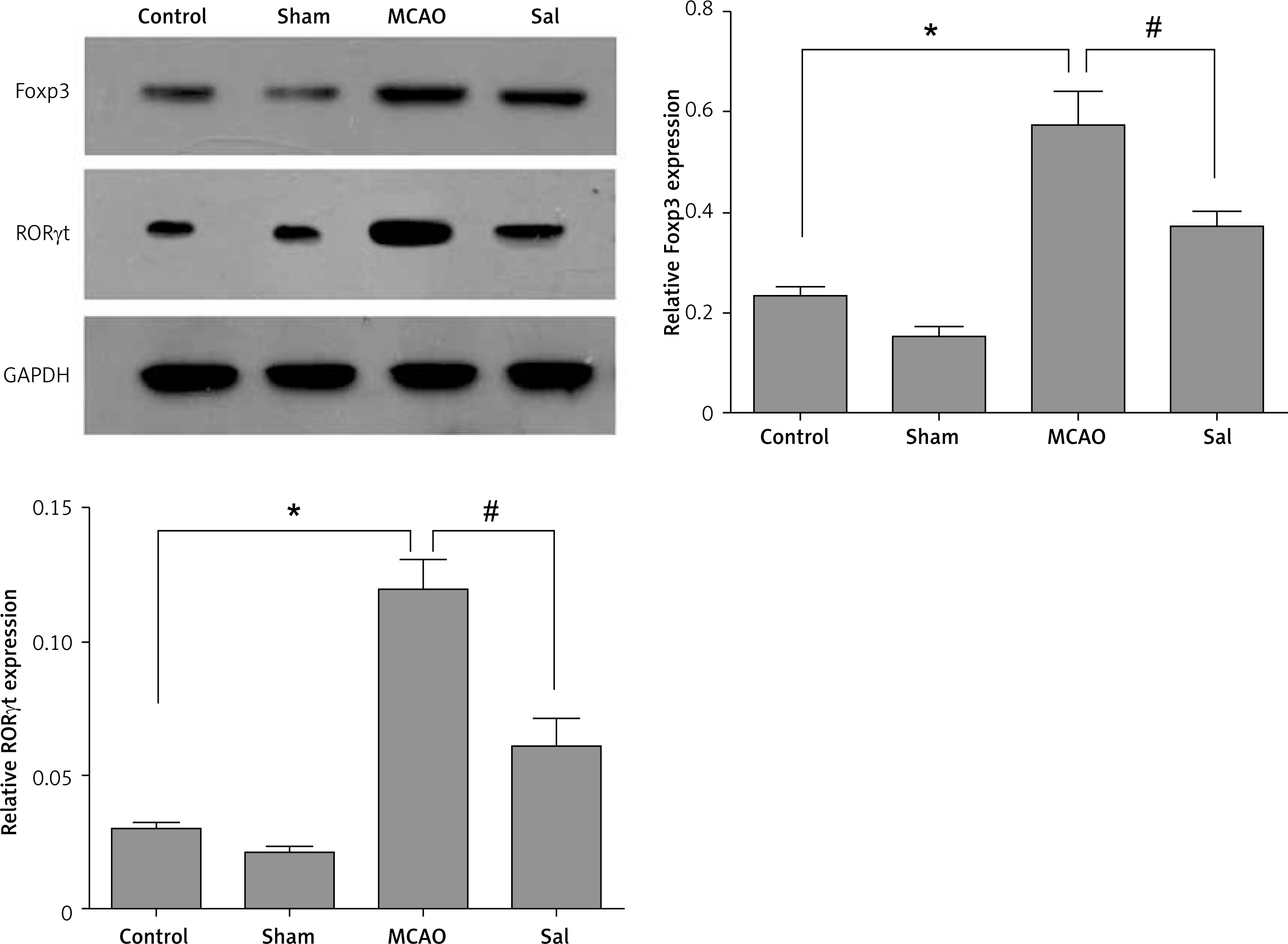
Sal can maintain the balance between Th17 and Treg cells
Western blot results showed that the expression levels of RORγt and Foxp3 protein in the brain tissue of the MCAO group were significantly higher compared with the control group (Figure 3), while the Sal group was opposite. These results suggest that MCAO operation can induce expression of the transcription factor RORγt and Foxp3 protein in the rat neural system, and the changes in the expression of RORγt and Foxp3 protein directly lead to the change of the balance between Th17 and Treg cells. Also, Sal treatment can alleviate this trend.
Figure 3
Expression of Foxp3 and RORγt was detected with Western blot in rats’ brain tissue
Compared with the control group, *p < 0.05; compared with MCAO group, #p < 0.05.
As shown in Figure 4, the results of flow cytometry showed that MCAO could induce changes in the proportion of Th17 and Treg cells in the peripheral blood of rats, and Sal treatment can alleviate this trend.
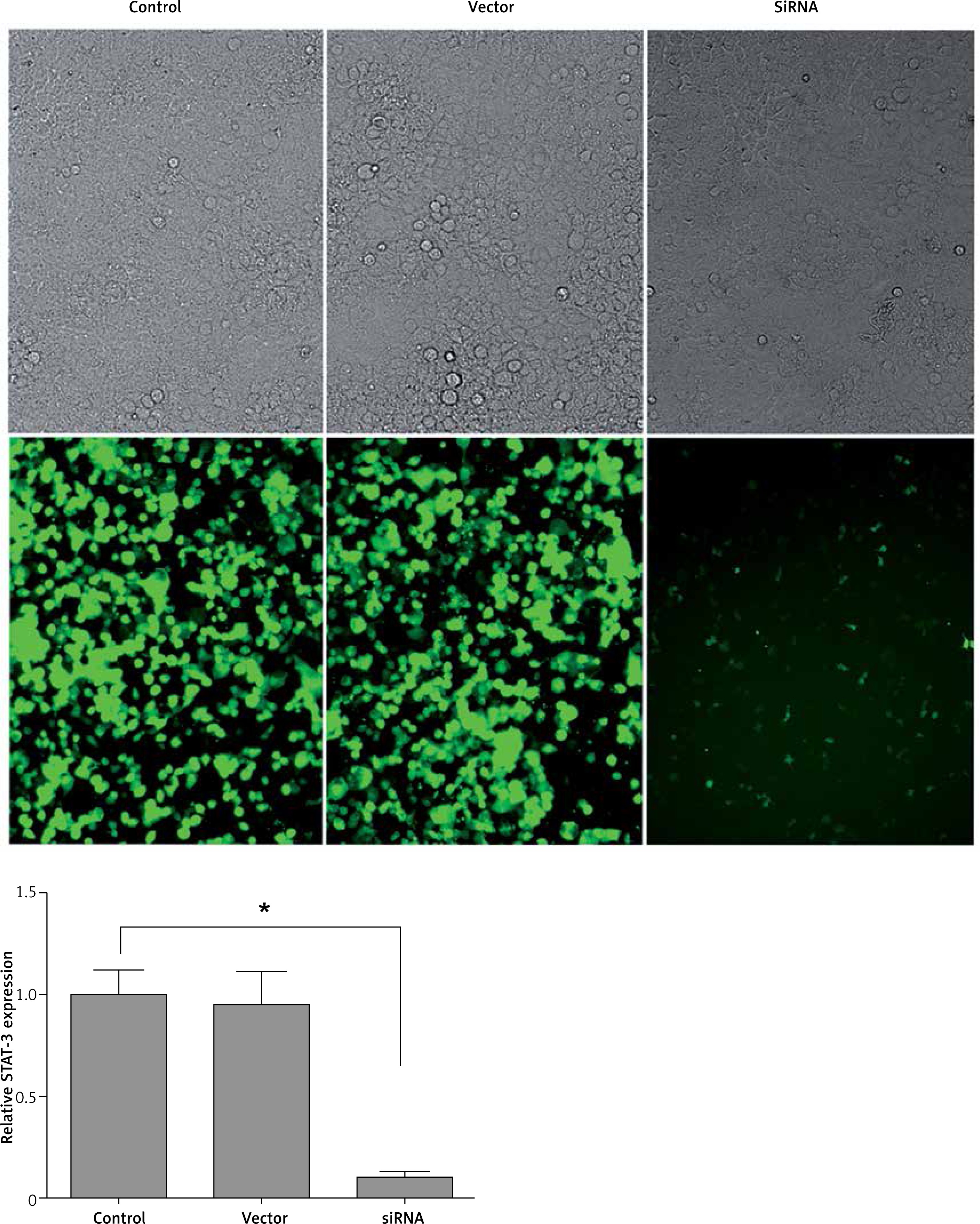
Successful construction of STAT-3 low expression cell line
The initial T cells were induced by IL-6 and TGF-β, and then Thl7/ Treg cells were obtained. The expression of transcription factor STAT-3 was downregulated by siRNA. The results showed that the expression of STAT-3 gene in T cells was inhibited successfully (Figure 5).
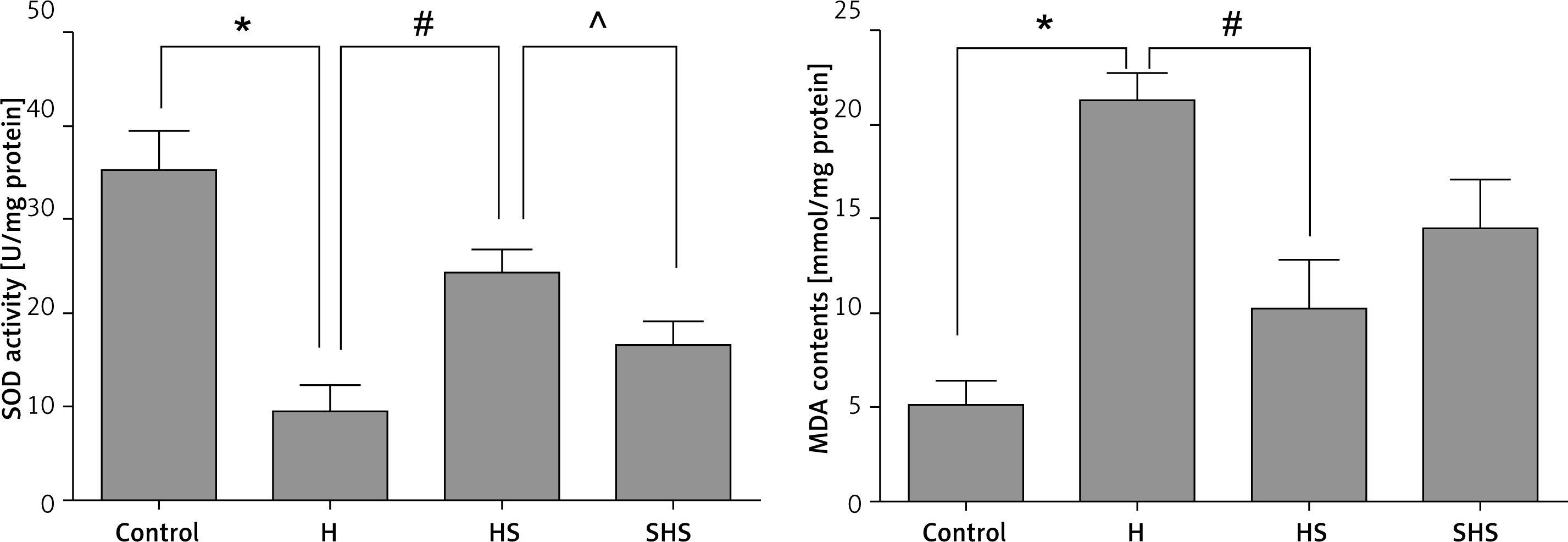
Sal can reduce the oxidative stress of T cells mediated by STAT-3
As shown in Figure 6, the results showed that Sal treatment could significantly increase the activity of SOD (Figure 6 A) and reduce the content of MDA (Figure 6 B). While the expression of STAT-3 was inhibited, the effect of Sal was inhibited as well. These results showed that Sal probably can reduce the oxidative stress of T cells, which may be mediated by STAT-3.
Figure 6
Assessment of oxidative states using assays of SOD and MDA in T cells after STAT-3 gene silencing
H – hypoxic condition, HS – hypoxic condition plus Sal, SHS – hypoxic condition plus Sal after STAT-3 gene silencing. Compared with the control group, *p < 0.05; compared with H group, #p < 0.05; compared with HS group, ^p < 0.05.
Sal can reduce the apoptosis of T cells and induce proliferation mediated by STAT-3
As shown in Figures 7 A and B, hypoxia treatment could significantly increase the apoptosis of T cells, and Sal treatment could significantly reduce the apoptosis of T cells after hypoxia treatment, while expression of the STAT-3 gene in T cells was inhibited, and Sal treatment lost its effect.
Figure 7
Apoptosis and proliferation of T cells after STAT-3 gene silencing. A, B – Apoptosis results of T cells treated under hypoxic conditions, Sal and STAT-3 gene silencing. C – MTT assays were performed to determine the proliferation in T cells
H – hypoxic condition, HS – hypoxic condition plus Sal, SHS – hypoxic condition plus Sal after STAT-3 gene silencing. Compared with the control group, *p < 0.05; compared with H group, #p < 0.05; compared with HS group, ^p < 0.05.
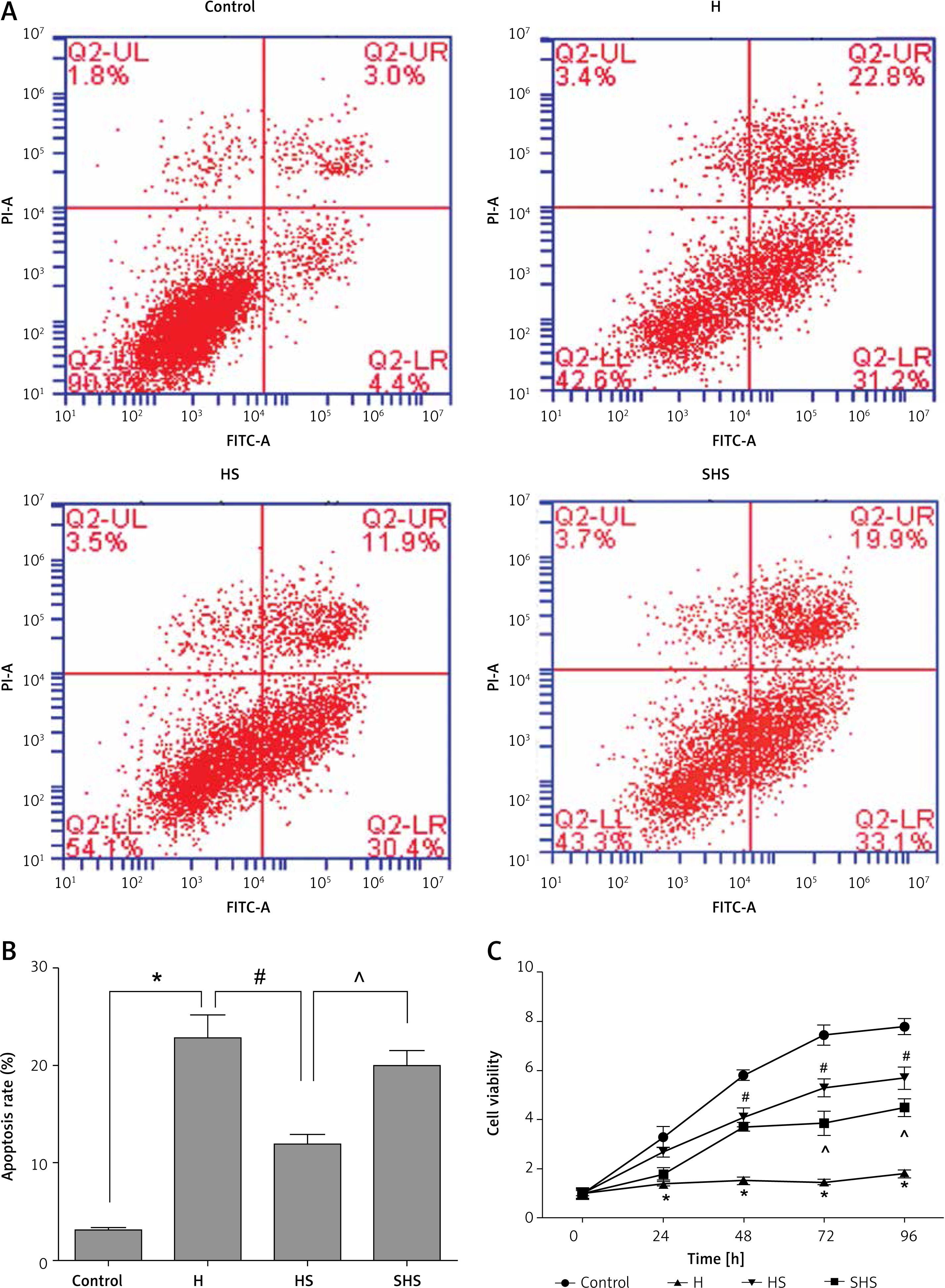
As shown in Figure 7 C, hypoxic treatment can significantly inhibit the activity of T cells, but after Sal treatment, cell viability increased significantly. However, it is interesting to note that after STAT-3 gene expression was inhibited, the role of Sal was also inhibited. These data indicate that Sal can promote the growth of T cells after anoxia treatment, which may be mediated by STAT-3.
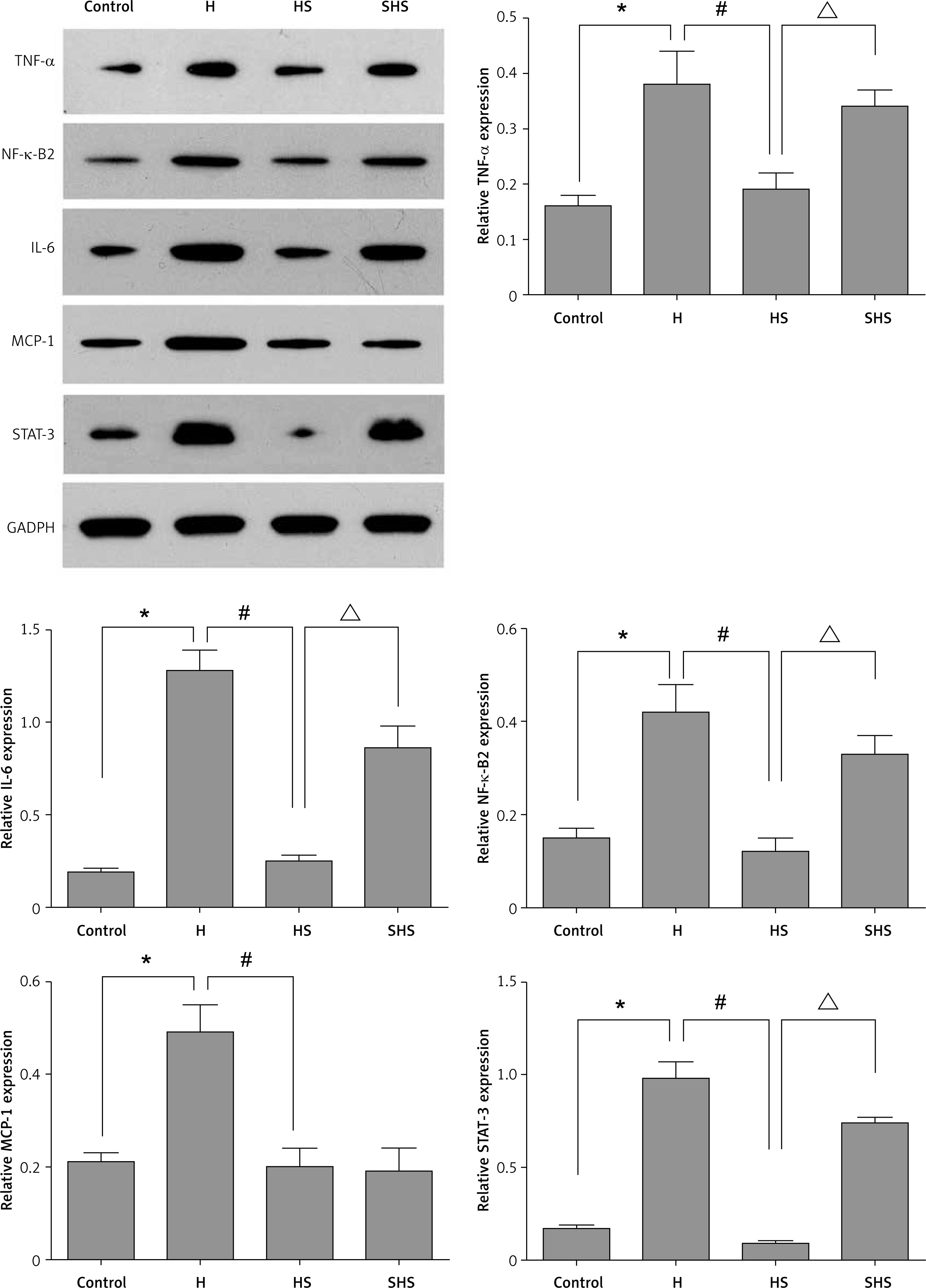
Sal can inhibit the expression of IL-6, TNF-α, MCP-1 and NF-κ-B2 protein by targeting STAT-3
Western blot was used to detect the expression of IL-6, TNF-α, MCP-1, STAT-3 and NF-κ-B2. The results showed that (Figure 8) Sal could significantly inhibit the expression of IL-6, TNF-α, MCP-1, STAT-3 and NF-κ-B2 protein in T cells after hypoxia treatment. However, this trend was offset by the knockdown of the STAT-3 gene. These data further support our hypothesis that Sal promotes ischemic tolerance, and inhibits apoptosis and oxidative stress by targeting STAT-3.
Figure 8
Expression of IL-6, TNF-α, MCP-1, STAT-3 and NF-κ-B2 with STAT-3 gene silencing in T cells
H – hypoxic condition, HS – hypoxic condition plus Sal, SHS – hypoxic condition plus Sal after STAT-3 gene silencing. Compared with the control group, *p < 0.05; compared with H group, #p < 0.05; compared with HS group, △p < 0.05.
Discussion
Salidroside has the effects of anti-hypoxia, anti-inflammatory, anti-apoptosis, anti-cancer and so on [24, 25]. In this study, we detected the effects of salidroside on oxidative stress in the brain tissue of rats. The results showed that Sal could reduce the neurologic deficit in MCAO rats and reduce the infarct area caused by local ischemia in rats. Cerebral ischemia can significantly induce apoptosis of brain cells. This conclusion has been further confirmed in our experiment. But in this study, we also found that the Sal treatment can alleviate this trend. MDA level and SOD activity indirectly reflected oxidative stress. In vitro data showed that Sal could reduce MDA content and enhance SOD activity in T cells. In addition, our results indicate that Sal treatment can significantly inhibit the apoptosis of ischemic brain cells. These results indicate that Sal has significant protective effects of brain injury induced by ischemia in rats. But the mechanism is not yet clear.
The Th17 cell is a CD4+ cell that can secrete IL-17 specifically under the regulation of nuclear factor ROR-γt. IL-17 can promote the secretion of IL-6, TNF-α and many other proinflammatory cytokines in the antigen presenting cells, and raise the concentration of granulocytes to the inflammatory sites, and participate in and expand the host inflammatory response [26]. Treg cells are also known as regulatory T cells. Cytokines TGF-β and IL-2 play a key role in the differentiation of Treg cells. TGF-β is the first discovered cytokine that can induce peripheral differentiation of Treg cells [27, 28]. As an important T cell growth factor, IL-2 plays a central role in thymic differentiation of Treg cells. IL-2 can activate the JAK-STAT5 pathway [29]. STAT5 can directly bind to the Foxp3 gene to promote Foxp3 gene transcription, thereby promoting Foxp3 expression and inducing Treg cell differentiation [30]. The differentiation and regulation of Thl7 and Treg cells are completed under their respective inducible factors and cell pathways, and the differentiation process shows a complex and fine network relationship [31]. The subgroups are resistant and balanced to each other through their respective transcription factors and cytokines. It can be seen that there is a balance between Thl7 and Treg cell subsets in the normal state of the body. Once the balance is destroyed, it will lead to the occurrence of immune diseases [32]. Maintaining Thl7/Treg cell balance is of great importance to the immune homeostasis and the treatment of diseases. The balance of ROR-γt and Foxp3 is an important way to maintain the balance of Thl7/Treg cells. Therefore, in this study, we detected the expression level of ROR-γt and Foxp3 protein in the brain tissue of MCAO rats by Western blot. The numbers of Th17 and Treg cells in peripheral blood of each group were detected by flow cytometry. Our experimental data showed that the expression of ROR-γt and Foxp3 protein can be significantly induced by cerebral ischemia in rats, and the Sal treatment can reverse this result. The number of Th17 and Treg cells in the peripheral blood of rats was detected by flow cytometry. The results showed that the MCAO operation could induce a change of Th17 and Treg cells in the peripheral blood of rats, which resulted in the imbalance of Th17 and Treg cells, but Sal could relieve the ratio of Th17 and Treg cells in the peripheral blood of the rats. In conclusion, the protective effect of Sal on brain injury induced by ischemia in rats may be achieved by regulating the proportion of Th17 and Treg cells. It has been reported that the dynamic balance of transcription factor STAT3/STAT5 can represent the balance between Thl7/Treg cells [33]. However, the relationship between STAT3/STAT5 and ROR-γt/Foxp3 is not clear. In this study, STAT3 was used as a further research object, and specific siRNA was used to inhibit the expression of STAT3 in T cells. The results showed that Sal could significantly reduce the apoptosis of T cells after hypoxia treatment, and the downregulation of STAT3 expression could inhibit the effect of Sal. This result suggests that the protective effect of Sal to hypoxia may be mediated by STAT3. In addition, we detected the proliferation characteristics of T cells. The results showed that Sal could promote the proliferation of hypoxic cells, while the knockdown of STAT3 could counteract the effect of Sal. Sal can significantly inhibit the expression of T cells IL-6, TNF-α, MCP-1, STAT-3 and NF-κ-B2 protein after hypoxia treatment. However, this state can be reversed by the knockdown of the STAT-3 gene. Studies have shown that STAT-5 can reduce the expression of RORγt and antagonize the differentiation and survival of Thl7 cells by competitive inhibition of STAT-3 [34]. However, in this study, we did not knock down the expression of the STAT-5 gene in T cells, and thus failed to verify the relationship between STAT-3/STAT-5 in the process of brain damage induced by Sal reduction by hypoxia. These studies will need further exploration in the future.
In conclusion, salidroside can reduce the neurological score of MCAO model rats, reduce the volume of cerebral infarction and have neuroprotective effects. This effect may be related to inhibition of the expression of IL-6, TNF-α, MCP-1, STAT-3 and NF-κ-B2. STAT-3 plays an important role in this process.