Introduction
Pre-eclampsia (PE) is a leading cause of maternal and morbidity worldwide [1]. This disease, which is characterized by proteinuria and hypertension, usually occurs at about 32 weeks of gestation [2]. It is estimated that approximately 2–8% of pregnant women suffer from PE [3, 4]. Several risk factors contribute to the pathogenesis of PE, including pregnancy age, times, hypertension, anemia, overweight/obesity, urinary tract infection, and hereditary factors [5, 6]. Despite the progress in the diagnosis and treatments, the current interventions are insufficient to achieve improving effects on the prevention and treatment of PE, as well as its complications [7]. Therefore, it is important to explore the mechanism of PE for further understanding.
A recent study revealed that failure in the vascular remodeling of the maternal spiral arteries is the main pathogenesis of PE, which leads to hypoperfusion of the placenta [8]. Specifically, trophoblast invasion is the one of the most important contributors to failure in vascular remodeling [9]. Hunkapiller et al. documented that the Notch signaling pathway plays a critical role in the endovascular invasion of the trophoblasts [10]. Since micoRNAs (miRNAs) have been demonstrated as therapeutic targets, they could be used as biomarkers in various diseases [11, 12]. Moreover, they are also reported to play critical roles in the regulation of trophoblast invasion, which participates in the pathogenesis of PE. Li et al. revealed that miR-29b promotes PE via regulating the apoptosis and invasion of trophoblasts [13]. Meanwhile, miR-16 suppresses the proliferation of mesenchymal stem cells in severe PE [14], miR-126 is downregulated in placentas of PE patients, and miR-126 correlates with the decreased expression of vascular endothelial growth factor (VEGF) [15]. Therefore, miRNAs might play critical roles in the regulation of PE pathogenesis.
In order to further explore the potential mechanism of miRNAs in PE, the bio-function of miR-210 and its downstream target were investigated, as well as the potentially involved pathway, so that some useful information could be provided for the diagnosis and treatment of PE.
Material and methods
Cell culture
The human placental choriocarcinoma cell line JAR was purchased from American Type Culture Collection (ATCC, Manassas, VA, USA) to explore the functions of trophoblastic cells. JAR cells were maintained in 1640 medium supplemented with 10% fetal bovine serum, 6 mM glutamine, 20 mM HEPES, 100 μg streptomycin and 100 IU penicillin at 37°C with a humidified atmosphere of 5% CO2. For hypoxia treatment, cells were maintained at 37°C with a humidified atmosphere of 5% CO2, 2% O2 and balanced N2.
Cell transfection
For knockdown of PTPN2 in JAR cells, short hairpin RNA targeting PTPN2 (sequence: GAAGATGTGAAGTCGTATTAT) or a control (sequence: 5′-AAAGAAATGTCCTACCTTCT-3′) was inserted into the plKO.1 vector to construct the shPTPN2 or control plasmid. Then, JAR cells were seeded in a 6-well plate with complete medium, and miR-210 inhibitor (purchased from GE Health Dharmacon, Waltham, MA, USA), miR-210 mimic (purchased from GE Health Dharmacon, USA), si-PTPN2 and si-PTPN2 control were respectively transfected into JAR cells using Lipofectamine 2000 (Invitrogen, USA) following the manufacturer’s protocol on the second day. After transfection for more than 48 h, cells were used for the following experiments.
MTT assay
The JAR cells with different transfections were seeded on a 96-well plate. After culture for 24 and 48 h, 200 μl of 5 mg/ml MTT (Sigma, St Louis, MO, USA) was added into each well at 37°C for 4 h. Then the medium was discarded and replaced with dimethyl sulfoxide. After shaking for 10 min, the absorbance of each well at 540 nm was determined using a microtiter plate reader.
Matrigel invasion assay
The JAR cells (5 × 105) with different expression levels of miR-210 or PTPN2 were plated in 1640 medium without FBS in the upper chamber of a transwell (Corning, Somerset, NJ, USA) coated with Matrigel (BD, MA, USA). Complete growth medium was added to the lower chambers. Then the plate was incubated in standard culture conditions (21% O2) or hypoxic conditions (2% O2) at 37°C for 72 h. Following this, cells on the upper chamber were removed with a cotton swab, and cells on the underside were stained with hematoxylin (Sigma). For each sample, 5 views were randomly selected and quantified, and the mean value was computed. Each experiment was performed in triplicate, and the mean value was calculated as the final result.
Transwell assay
Similarly, for the Matrigel assay, 5 × 105 JAR cells with different expression levels of miR-210 or PTPN2 were separately seeded in 1640 medium without FBS in the upper chamber of a transwell (Corning). Then, the lower chamber was filled with complete growth medium (1640 medium with FBS). Next, the plate was cultured in standard conditions (21% O2) or hypoxic conditions (2% O2) at 37°C for 72 h. Subsequently, cells on the upper chamber were scraped with a cotton swab, and on the other side were stained with hematoxylin (Sigma). For each sample, 5 views were randomly selected and qualified, and the mean value was obtained as the final result of the sample. Each experiment was performed in triplicate, and the mean value was calculated as the final result.
Quantitative real-time PCR (qRT-PCR)
After culture or treatment, medium of JAR cells was completely removed, and 1 ml of Trizol agent (Invitrogen) was added to isolate total RNA according to the manufacturer’s protocol. After quantification using a NanoDrop spectrophotometer (NanoDrop Technologies, Oxfordshire, UK), 2 μg of total RNA was used for mRNA reverse transcription using the high-capacity cDNA Reverse Transcription kit (Applied Biosystems, lnc., Foster City, CA, USA) according to its specific instructions. Meanwhile, 2 μg of total RNA was used for miRNA reverse transcription using the TaqMan miRNA Reverse Transcription Kit (Applied Biosystems, lnc., Foster City, CA, USA). Then, the expression levels of the gene and miRNA were determined on an ABI PRISM 7900HT PCR system (Applied Biosystems, Foster City, CA, USA) with the following system: for the quantification, GAPDH was used to normalize mRNA, and U6 was used to normalize miRNA. Primers of miR-210 and PTPN2 were designed as follows: miR-210-forward, 5′-GTGCAGGGTCCGAGGT-3′, and miR-210-reverse: 5′-CTGTGCGTGTGACAGCGGCTGA-3′; U6-forward, 5′-CTCGCTTCGGCAGCACA-3′, and U6-reverse, 5′-AACGCTTCACGAATTTGCGT-3′; PTPN2-forward, 5′-TTCCTCTGAACCCCAAACTG-3′ and PTPN2-reverse, 5′-GCCTCCAAAAACAAATCCTG-3′; and GAPDH-forward, 5′-CGAGATCCCTCCAAAATCAA-3′ and GAPDH-reverse, 5′-TGTGGTCATGAGTCCTTCCA-3′. Then, the fold changes of genes and miRNA were calculated using the 2–ΔΔCt method [16]. Each sample was analyzed in triplicate, and the mean value was set as the final expression value.
Western blotting
Protein lysates from each cell sample were collected, quantified with the Bradford assay, and boiled with loading buffer for 10 min. A total of 10 μg of protein was used to run in 10% or 12% SDS-PAGE gel, and transferred electrophoretically to a PVDF membrane using a standard method. After blocking with skimmed milk, membrane was incubated with rabbit anti-PTPN2 (1 : 1000), rabbit anti-PDGFR (1 : 2000), p-PDGFR (1 : 1000), Akt (1 : 3000), p-Akt (1 : 2000), Erk (1 : 3000), p-Erk (1 : 3000), or GAPDH (1 : 5000) purchased from Cell Signaling Technology (Denvers, MA, USA), then incubated with secondary rabbit antibodies (1 : 10000, Cell Signaling Technology), and visualized using the enhanced chemical luminescence method.
Statistical analysis
In the current study, SPSS 20.0 software (IBM, SPSS, Chicago, IL, USA) was used to perform statistical analysis. Continuous variables were presented as mean ± standard deviation. Comparisons between groups were estimated using Student’s t-test, and p < 0.05 was considered statistically significant.
Results
Hypoxia induces expression of miR-210 in JAR cells
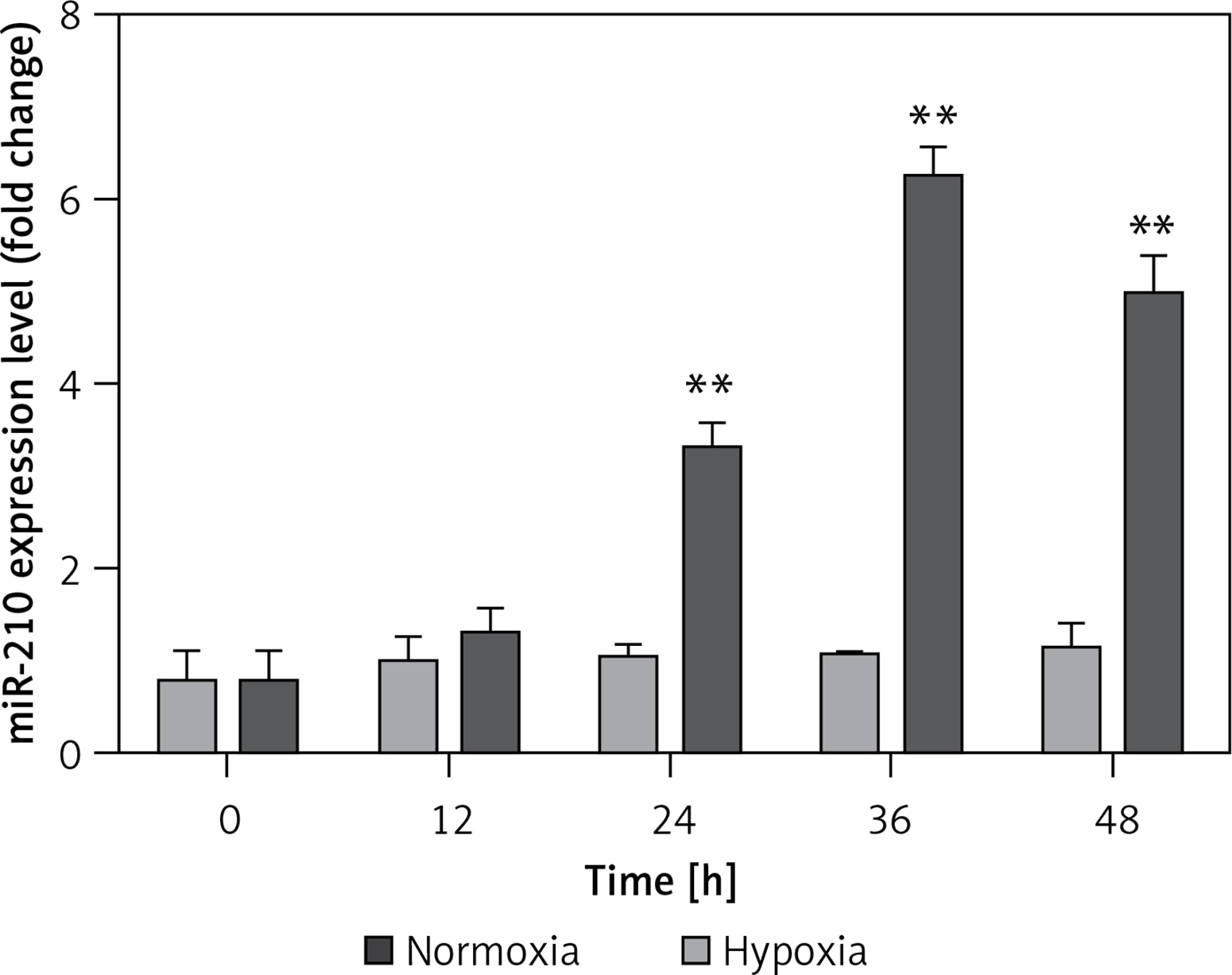
To investigate the bio-function of miR-210, JAR cells were cultured in standard conditions with 21% O2 and hypoxic conditions with 2% O2. The results showed that the mRNA level of miR-210 significantly increased in a time-dependent manner within 48 h under hypoxic conditions compared with the control group (Figure 1), indicating that miR-210 was involved in the response to hypoxia.
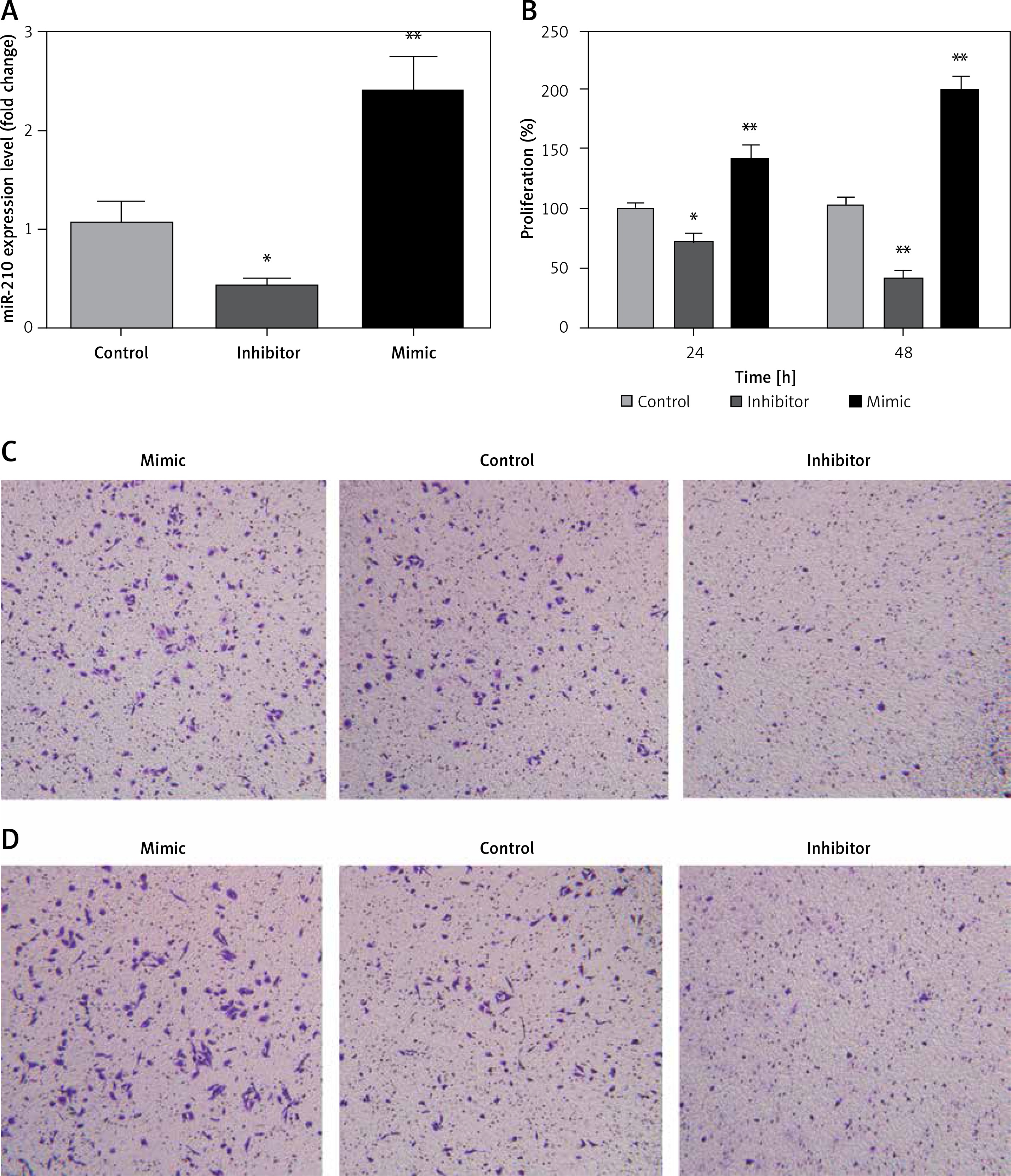
miR-210 promotes the cell viability, migration and invasion of JAR cells
For further analysis, miR-210 control, mimic and inhibitor were separately transfected into JAR cells, and the efficacy was determined using RT-PCR (Figure 2 A). Then, cell viability was measured using MTT, and the result showed that compared with the control group, miR-210 mimic significantly increased the viability of JAR cells, while miR-210 inhibitor markedly decreased cell viability of JAR cells (Figure 2 B). Similar tendencies were also identified in the migration and invasion of JAR cells (Figures 2 C, D). These findings indicated that miR-210 could increase cell viability and promote migration and invasion of JAR cells.
Figure 2
Effect of miR-210 on the proliferation, migration, and invasion of JAR cells. A – Expression of miR-210 determined using real-time PCR, B – cell proliferation determined using MTT, C – cell migration assessed using transwell assay, D – cell invasion estimated using Matrigel assay. Compared with the control group, **p < 0.01
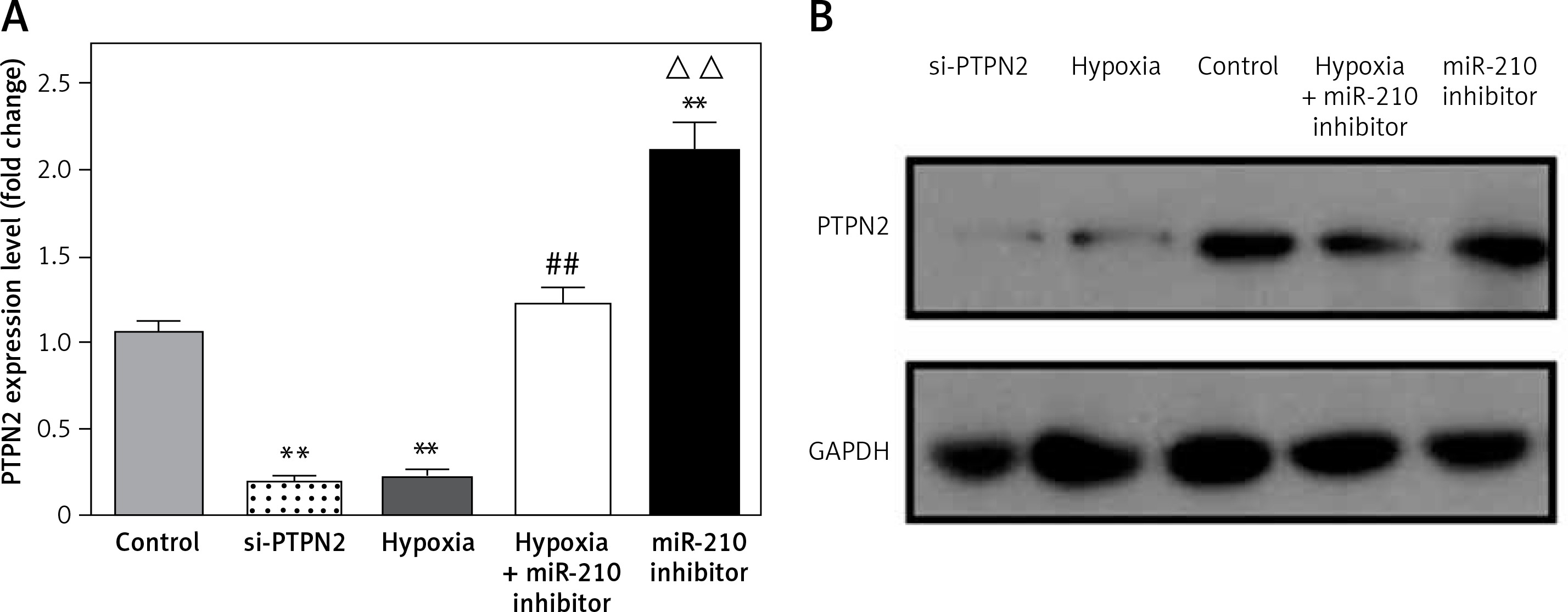
PTPN2 is a downstream target of miR-210
PTPN2 has been reported to be a target of miR-210 [17]. In the present study, shRNA was used to inhibit the expression of PTPN2 in JAR cells. Meanwhile, JAR cells were transfected with miR-210 inhibitor or treated with hypoxia to assess the correlation between miR-210 and PTPN2. Then, the expression levels of PTPN2 were measured using RT-PCR and Western blotting. As shown in Figure 3, both si-PTPN2 and hypoxia significantly decreased the expression of PTPN2, while miR-210 inhibitor prevented this elevation. These findings indicated that PTPN2 might be a target of miR-210, and inhibiting the expression of miR-210 could significantly increase the expression of PTPN2.
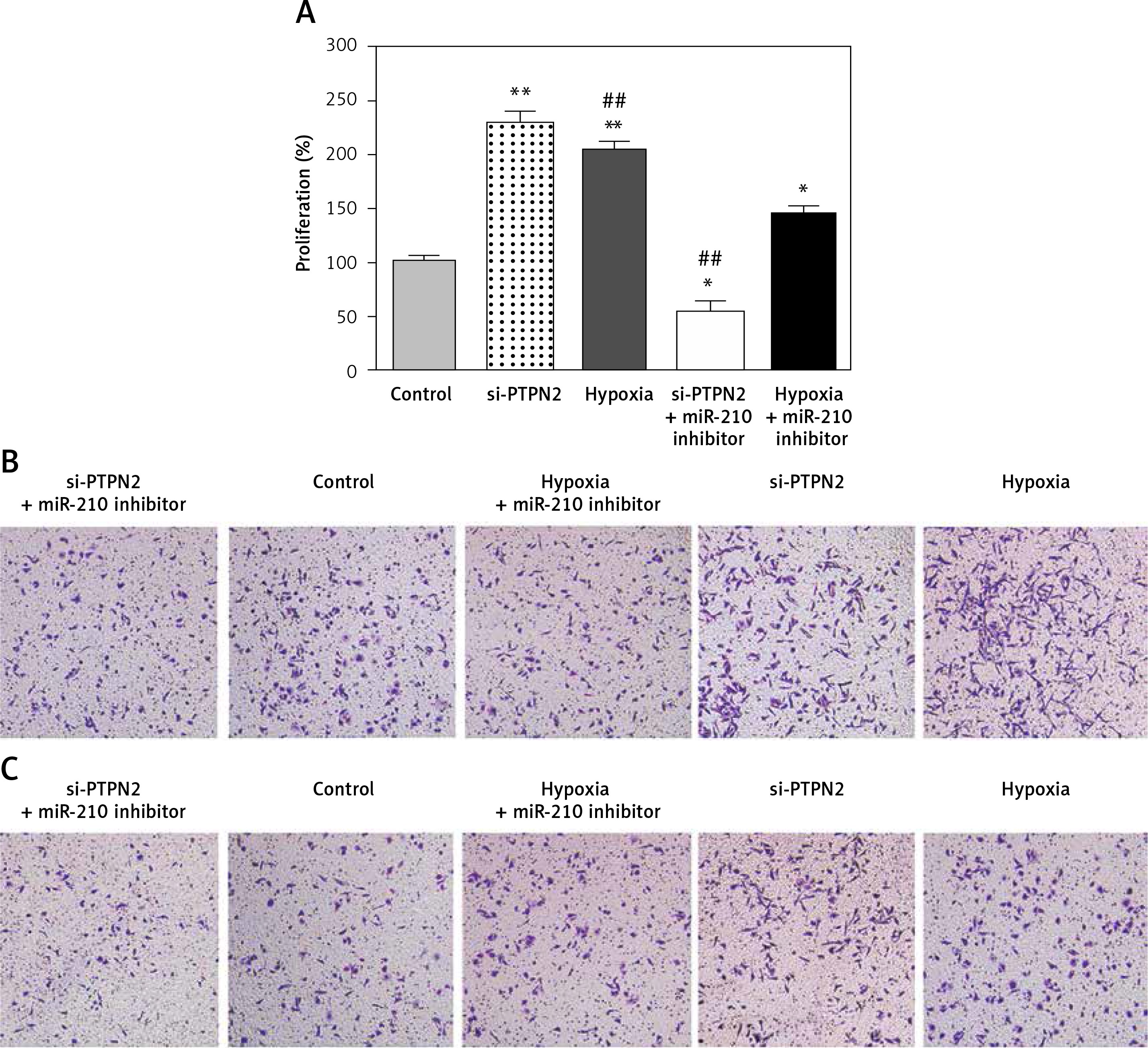
PTPN2 suppresses the proliferation, migration and invasion of JAR cells
Effects of PTPN2 on cell proliferation, migration, and invasion were investigated. Both si-PTPN2 and hypoxia significantly increased cell proliferation, and promoted cell migration and invasion of JAR cells, while miR-210 inhibitor reversed these effects of hypoxia and si-PTPN2 (Figure 4).
Figure 4
Effect of si-PTPN2 and hypoxia on the proliferation, migration, and invasion of JAR cells. A – cell proliferation determined using MTT, B – cell migration assessed using transwell assay, C – cell invasion estimated using Matrigel assay
Compared with the control group, *p < 0.05 and **p < 0.01; compared with hypoxia + miR-210 inhibitor, ##p < 0.01.
Pathway of miR-210-PTPN2 involvement
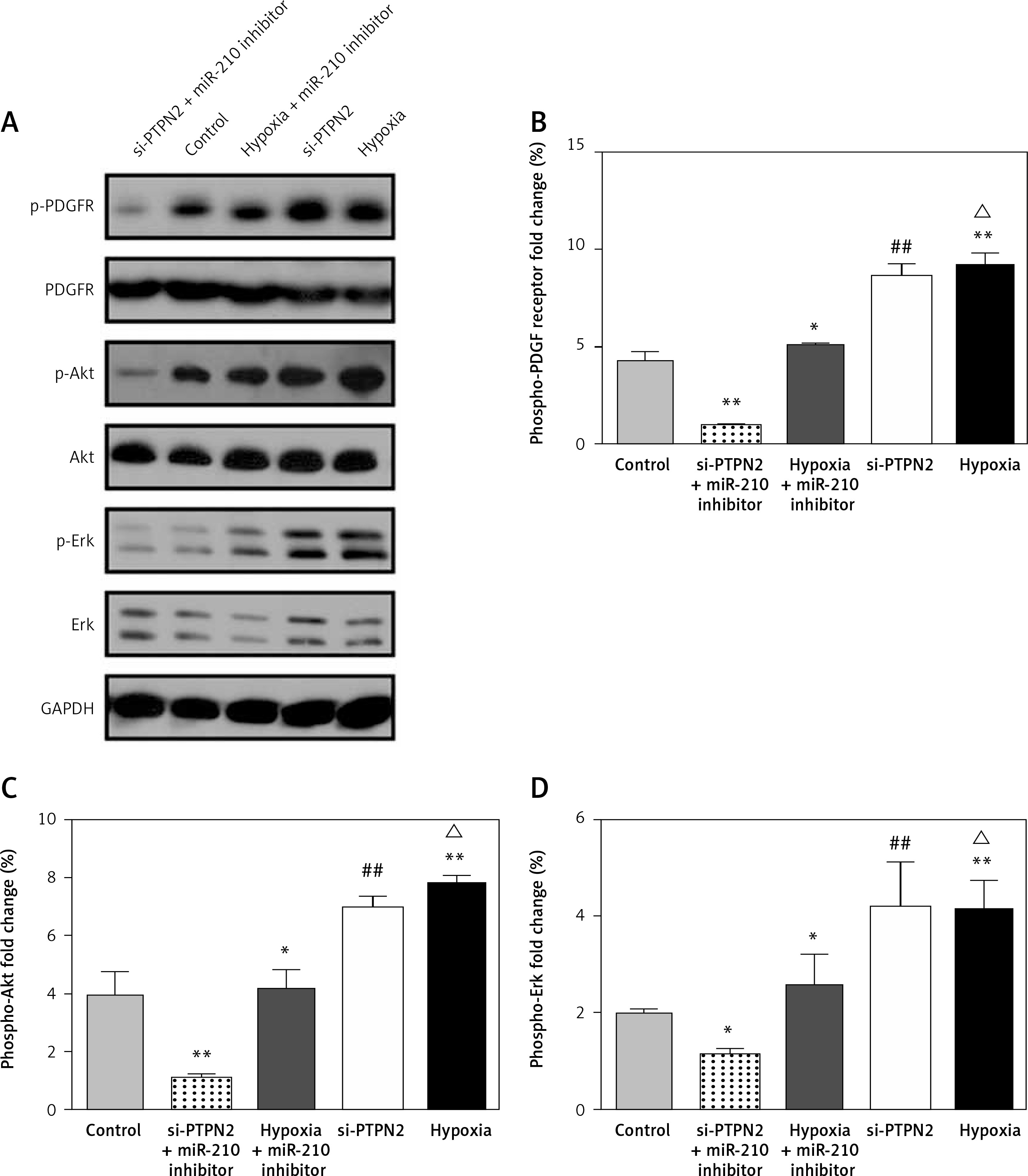
To explore the potential signaling pathway of miR-210-PTPN2 involved in the current study, the expression and phosphorylation levels of PDGFR, Akt, and ERK were determined by Western blotting (Figure 5 A). The quantification of phosphorylation showed that si-PTPN2 and hypoxia treatment significantly increased the phosphorylation levels of PDGFR (Figure 5 B), Akt (Figure 5 C), and ERK (Figure 5 D) compared with the control group, and miR-210 reversed these outcomes, with no significant changes in these levels. These findings indicated that miR-210-PTPN2 might regulate the cell proliferation, migration and invasion via the PDGFR-Akt signaling pathway.
Figure 5
Expression and phosphorylation variations of PDGFR, Akt and Erk. A – Variations determined using western blotting, B – quantification of phosphorylation change of PDGFR, C – quantification of phosphorylation change of Akt, D – quantification of phosphorylation change of Erk
Compared with the control group, *p < 0.05 and **p < 0.01; compared with hypoxia + miR-210 inhibitor, ##p < 0.01.
Discussion
In the current study, the bio-function of miR-210 and its potential target were revealed in JAR cells. After investigation, miR-210 expression increased significantly under hypoxia. Overexpression of miR-210 obviously promoted the cell proliferation, metastasis and invasion of JAR cells, and PTPN2, a target of miR-210, significantly attenuated these elevations via the PDGFR-Akt signaling pathway.
Among hypoxia-regulated miRNA, miR-210 has been reported to be involved in several diseases. Grosso et al. documented that miR-210 promotes a hypoxic phenotype and radioresistance of human lung cancer cell lines [18]. Meanwhile, Kai et al. reported that miR-210 is also involved in the regulation of hepatocellular carcinoma via HIF-1α and HIF-3α [19]. Moreover, McCormick et al. demonstrated that miR-210 is a target of HIF-1 and HIF-2, and is closely correlated with the prognosis of patients with renal cancer [20]. Based on these findings, it can be supposed that miR-210 plays a critical role in the hypoxia response. Zhang et al. revealed that expression of miR-210 could be up-regulated under hypoxia by NF-κB transcriptional factor p50 in PE [21]. Both Gan et al. and Nikuei et al. identified that miR-210 and miR-155 might be potential diagnostic markers for PE [22, 23]. All these findings imply that miR-210 also might play a crucial role in the pathogenesis of PE. However, the underlying mechanism remains unclear. Complying with the previously published studies, the present study identifies PE with a significant increase in the expression of miR-210 [21, 24, 25]. Further analyses showed that the overexpression of miR-210 could significantly promote cell proliferation, migration, and invasion, while miR-210 inhibitor could significantly inhibit these behaviors of JAR cells. These findings indicate that hypoxia induces the elevation of miR-210 and promotes the invasion of trophoblast cells, as well as the failure of arterial remodeling, leading to the occurrence and development of PE.
As an important member of the protein-tyrosine phosphatase (PTP) family, PTPN2 is involved in the regulation of T-cells and its related biological process. Wiede et al. reported that deficiency of PTPN2 enhanced the response of B cells and T cells, as well as systemic inflammation and autoimmunity [26]. Meanwhile, another study reported that PTPN2 could suppress the proliferation of T-cells [27]. Moreover, Spalinger et al. documented that PTPN2 could regulate the differentiation of CD4+ T cells, and inhibit inflammation in the intestine [28]. Recently, Kim et al. demonstrated that miR-210 promotes the proliferation and migration of stem cells derived from adipose tissue via downregulating PTPN2 [29]. Thus, the bio-function of PTPN2 was uncovered in the current study. The analytical results showed that expression of PTPN2 could be decreased by hypoxia, and miR-210 inhibitor could inhibit this downregulation, which was also identified by Adel et al. in PE [30]. Moreover, analyses of cell behaviors showed that si-PTPN2 could induce the proliferation, migration, and invasion of JAR cells, indicating that PTPN2 performed a negative role in the development of PE. However, miR-210 inhibitor could obviously suppress the effect of si-PTPN2 or hypoxia on JAR cells, indicating that PTPN2 might be a downstream target of miR-210 in the pathogenesis of PE. In addition, PDGFR is reported to be common substrate for PTPN2 [31]. Then, the protein and phosphorylation levels of PDGFR were assessed, as well as Akt and Erk, which are involved in the same signaling pathway. The results showed that the phosphorylation levels of PDGFR, Akt, and Erk were significantly increased by hypoxia treatment and si-PTPN2, but this was prevented by miR-210 inhibitor, indicating miR-210-PTPN2 might regulate the development of PE via the PDGFR-Akt signaling pathway. However, the detailed mechanism still needs to be further explored.
Taken together, this study documented that miR-210 plays a critical role in the development of PE via targeting PTPN2, which could inhibit the process of PE via the PDGFR-Akt pathway. Hence, both miR-210 and PTPN2 could serve as potential biomarkers and therapeutic targets of PE. There are some limitations that restrict the power of this study. First, the expression of miR-210 and PTPN2 was not examined in clinical samples due to the small sample size. Second, in vivo validation was not provided in this study owing to deficiency of ethic authorization. Last but not least, the in-depth mechanism of miR-210 in PDGFR-Akt pathway was not well established. Therefore, further exploration is still required to further reveal the mechanism of miR-210 and PTPN2 in PE. Despite these issues, this study provided some basic information on the process of PE. In conclusion, miR-210 expression could be significantly induced under hypoxia, but PTPN2 expression was obviously decreased under hypoxia, and these variations could significantly promote the proliferation, migration, and invasion of JAR cells. Downstream of miR-210, the effects of si-PTPN2 on JAR could be markedly suppressed by miR-210 inhibitor via the PDGFR-Akt signaling pathway. All of this evidence indicates that miR-210 and PTPN2 might act as potential biomarkers and therapeutic targets for the diagnosis and treatment of PE.