Introduction
Hepatocellular carcinoma (HCC) accounts for 90% of primary liver cancers and it represents the third most common cause of death from cancer worldwide, with an increasing incidence expected in the next decades [1]. The major risk factors are chronic viral hepatitis B and C (HBV and HCV), alcohol abuse, primary biliary cirrhosis, xenobiotics, diabetes, non-alcoholic fatty liver disease and genetic disorders such as hemochromatosis and α1-antitrypsin deficiency [2, 3]. To date, surgery remains the most effective treatment with curative potential. However, only about 10% to 20% of patients with HCC are currently eligible for surgical intervention [4]. Therefore, it is urgently required to find a sensitive biomarker to detect liver cancer at the early stage. Egypt has the highest prevalence of HCV in the world, estimated nationally at 14.7% [1], with genotype 4 being the most common [2]. Liver cancer development is thought to develop in a multi-step process requiring the accumulation of several structural and genomic alterations, thus affecting many different molecular pathways [5].
MicroRNAs (miRNAs) are an important class of small non-coding RNA molecules that have recently come to prominence as critical regulators in a wide array of mechanisms of cell physiology. There is increasing evidence that miRNAs may also have an important function in viral replication and may be used by host cells to control viral infection [6].
Cellular miRNAs may recognize viral RNAs and silence them [7]. It has been suggested that many of the miRNA changes that take place during hepatocarcinogenesis do so early; therefore, many changes that predispose to liver cancer have already taken place during liver cirrhosis and other premalignant lesions [8]. It was found that specific circulating miRNAs could be detected significantly earlier after liver injury. In 2006, it was first reported that down-regulation of miR-122 could be a potential biomarker for liver cancer in rodent and human HCC [9].
MiRNAs are not only found inside cells but they are also circulating in serum and other bodily fluids. The function of these extracellular circulating miRNAs is not well understood yet. They may be released due to tissue damage as in the case of miR-122 and miR-192, which is indicative of liver degeneration in mice in response to drug treatment [10]. They also act as mediators of cell-cell communication as seen in Epstein-Barr virus infected cells where miRNAs secreted by infected cells are taken up by monocyte-derived dendritic cells repressing certain target genes in the dendritic cells [11]. Changes in circulating miRNA profiles have been associated with various diseases. They may be used as potential biomarkers because of their stability in body fluids, sensitive detection by quantitative polymerase chain reaction (PCR), and non-invasiveness in obtaining samples of body fluids [12]. For example, serum miR-21, miR-122 and miR-223 were seen to be elevated in patients with HCC or chronic hepatitis, which means that these miRNAs could be used as diagnostic biomarkers for liver injury [13].
Although miRNA-based diagnostics has been successfully utilized to diagnose, monitor, and/or manage a wide range of neoplastic and non-neoplastic disorders, several knowledge-based and practical issues hamper their translation into routine clinical practice. These issues include lack of collaborative research efforts, miRNA sequence database errors, non-standardization of sample processing protocols, analytical variation, suboptimal RNA extraction methods, lack of an accurate and valid normalization protocol, non-reliability and non-reproducibility of many results, detection assays variability, and lack of cellular and clinical database resources [14].
Thus, the aim of the current work was to develop a non-invasive reliable diagnostic biomarker through a carefully designed study based on measuring the serum levels of different miRNAs in order to detect HCV genotype 4-induced HCC at the early stages of the disease.
Material and methods
This study involved 165 patients admitted to the Hepato-Gastroenterology Department, Theodor Bilharz Research Institute (TBRI), Giza, Egypt for evaluation of their HCV-related chronic liver disease. All had circulating anti-HCV genotype 4 antibodies, and no serologic evidence of co-infection with HBV. Patients were excluded from the study if they had parasitological, serological, histopathological, or ultrasonographic findings indicative of other etiologies of chronic liver disease (CLD) other than HCV, such as non-alcoholic steatohepatitis, autoimmune hepatitis, biliary disorders, malignancies other than HCC, bacterial infection, Schistosoma infection, or dual chronic hepatitis B and C infection.
All patients gave informed consent prior to inclusion in the study in conformance with the guidelines of the 1975 Declaration of Helsinki as reflected by approval of the institution’s human research ethics committee. All procedures were medically indicated for patient management. Clinically and biochemically healthy, roughly age- and gender-matched persons (n = 40) served as control subjects.
The patients were subjected to thorough clinical examination and were assessed by laboratory investigations: ultra-sonography and liver biopsy using ultrasound-guided Menghini needle. HCC diagnosis is made upon the presence of hepatic lesion suggestive of malignancy by abdominal ultrasonography and triphasic abdominal computed tomography (CT) scan and confirmed to be HCC by histologic assessment of liver biopsy.
Histopathologic study
Assessment of grade of inflammation and stage of fibrosis was carried out in 5 µm thick serial sections of formalin-fixed, paraffin-embedded blocks, stained with hematoxylin/eosin and Masson trichrome stains. The stage of hepatic fibrosis was determined according to the METAVIR scoring system (F0, F1, F2, F3 & F4). We collected the F0, F1, F2 & F3 in one group, the chronic hepatitis C (CHC) without cirrhosis and the F4 only in another group, the CHC with cirrhosis (LC) group.
Laboratory investigations
Liver function tests were done using commercially available kits. Hepatitis B markers were tested using enzyme immunoassay (EIA) kits (Axiom Diagnostics, Burstadt, Germany). Circulating anti-HCV (genotype 4) antibodies were detected using an EIA kit (Axiom Diagnostics, Burstadt, Germany), and the presence of HCV-RNA in patients’ sera was detected by real-time PCR using the Amplicor test (Roche Diagnostic Systems; Meylan, France). Meanwhile, serum AFP was tested using the Eurogenetics EIA kit (Tessenderlo, Belgium).
HCV detection
Viral RNA was extracted using the QIAamp Viral RNA kit (Qiagen, Valencia, CA) and stored at –80°C. The serum HCV-RNA level was measured using One-Step Real-Time PCR according to manufacturer’s protocol (Carlsbad, CA, USA).
Serum preparation and RNA extraction
For RNA isolation from serum, 250 µl of serum was homogenized in 750 µl of Trizol LS (Invitrogen, Carlsbad, CA, USA). Then 200 µl of chloroform was added to the sample and the mixed solution was centrifuged. After an additional chloroform extraction and precipitation with isopropanol, the pellet was washed twice by centrifugation with 70% ethanol. The RNA pellet was dried for 10 min at room temperature and dissolved in 30 µl of diethylpyrocarbonate (DEPC)-treated water. DNase treatment (Qiagen, Valencia, CA) was carried out to remove any contaminating DNA. The concentration and quality of RNA were measured by A260/280 ratio and checked by gel electrophoresis individually.
Reverse transcription (RT) and quantitative PCR (qPCR)
RT and qPCR kits made specifically for accurate miRNA analysis (Applied Biosystems, Foster City, CA, USA) were used. For real-time PCR, 1.33 µl of diluted RT products were mixed with 10 µl of 2 × TaqMan PCR master mixture, 1 µl of TaqMan MicroRNA assay and 7.67 µl of nuclease-free water in a final volume of 20 µl according to the manufacturer’s instructions. All reactions were run on the ABI 7300 (Applied Biosystems, USA) using the following conditions: 95°C for 10 min, followed by 40 cycles at 95°C for 15 s, and 60°C for 1 min. Real-time PCR was done in triplicate, including no-template controls. Relative expression of miRNA was calculated using the comparative cycle threshold (CT) (2–ΔΔ CT) method [15], with miRNA-39 as the endogenous control to normalize the data.
Statistical analysis
Due to the magnitude and range of relative miRNA expression levels observed, results data were log transformed for analysis. Data are presented as mean ± SE. There was no evidence against normality for the log transformed data as confirmed using the Kolmogorov-Smirnov test. Analysis of variance (ANOVA test) was used to evaluate expression differences of the chosen miRNAs between cases and controls.
Receiver operating characteristic (ROC) curves were constructed and the area under the curve (AUC) was calculated to evaluate the specificity and sensitivity of predicting cases and controls. All statistical tests were two-sided, and a probability level of p < 0.05 was considered to be statistically significant. Data analysis was done using SPSS 20.0 software (IBM Corp.).
Results
Demographic, laboratory investigations, and clinical features of the patients
Clinically and biochemically healthy, roughly age- and gender-matched individuals (n = 40) served as a control population for patients with HCV-related liver fibrosis, cirrhosis, and primary HCC. Table I summarizes the epidemiological data of control subjects and patients, i.e. number, age, and gender. The biochemical parameters, i.e. aspartate aminotransferase (ASP), alanine aminotransferase (ALT), albumin, alkaline phosphatase (ALP), prothrombin concentration, α-fetoprotein (AFP) and des-γ-carboxy prothrombin (DCP), were as expected, within the reference range for control subjects.
Table I
Demographic and laboratory data of all studied groups
| Variables | Controls (n = 40) | CHC with early fibrosis (F0 + F1 + F2 + F3) (n = 42) | CHC with cirrhosis (F4) (n = 45) | HCV-induced HCC (n = 38) |
|---|---|---|---|---|
| Age | 47.0 ±3.5 | 46.4 ±7.3 | 51.3 ±4.7 | 48.1 ±4.1 |
| Gender (M : F) | 3 : 1 | 3 : 2 | 7 : 5 | 6 : 4 |
| Liver function tests, mean ± SD: | ||||
| AST [U/l] | 21.32 ±1.07 | 46.14 ±4.77* | 50.84 ±5.00* | 76.40 ±7.34** |
| ALT [U/l] | 23.16 ±1.95 | 44.12 ±5.50* | 50.84 ±5.00* | 76.40 ±7.34** |
| ALP [U/l] | 189 ±41 | 331 ±36* | 336 ±48* | 420 ±33** |
| Albumin [g/dl] | 4.4 ±0.5 | 3.60 ±0.74 | 3.8 ±0.72 | 3.08 ±0.48 |
| Prothrombin concentration | 95.6 ±3.4 | 89.6 ±4.8 | 41.5 ±11.1** | 69.4 ±3.7* |
| AFP [IU/ml] | 3.12 ±0.08 | 8.86 ±0.11* | 10.11 ±0.11* | 55.18 ±0.44** |
| DCP [mAU/ml] | 30.42 ±0.70 | 121.49 ±0.59* | 123.62 ±0.38* | 456.52 ±0.66** |
Data are expressed as mean ± standard deviations (SD), CHC – chronic hepatitis C, HCC – hepatocellular carcinoma, HCV – hepatitis C virus, normal range for ALT and AST is up to 40 IU/l, normal range for ALP is up to 250 U/l, normal range for albumin is 3.5–5 g/dl, normal range for prothrombin concentration is 80–100%, normal range for AFP is 0.1–9.6 IU/ml, mAU/ml – milli-absorbance unit/ml, normal range for des-γ-carboxyprothrombin (DCP) ≤ 40 mAU/ml,
Serum HCV-RNA revealed that all patients were genotype 4. All the studied groups’ patients had increased ALT, AST, albumin, ALP, prothrombin concentration, AFP and DCP (p < 0.001) compared to the control group and the AFP and DCP serum levels were increased significantly in HCC patients compared to hepatitis C and cirrhosis patients (p < 0.001) (Table I).
Serum expression levels of miRNA-122a, miRNA-125a, miRNA-139, miRNA-145 and miRNA-199a
Serum levels of miRNA-122a, miRNA-125a, miRNA-139, miRNA-145 and miRNA-199a were significantly lower (p < 0.01) in HCC than in both CHC and LC groups. As a single marker, miRNA-122a had the highest sensitivity for HCC, followed by miRNA-199a, miRNA-145, miRNA-139 and miRNA-125a (Table II).
Table II
Real-time qPCR expression levels of microRNAs in serum of the different studied groups
| MiRNAs/groups | Control | CHC with early fibrosis (F0 + F1 + F2 + F3) (n = 42) | CHC with cirrhosis (F4) (n = 45) | HCV-induced HCC (n = 38) |
|---|---|---|---|---|
| miRNA-122a | 19.69 ±0.33 | 545.83 ±0.79** | 520.94 ±0.77** | 16.13 ±0.38a |
| miRNA-125a | 20.57 ±0.54 | 96.01 ±4.36** | 100.54 ±0.81** | 29.96 ±0.57** |
| miRNA-139 | 20.53 ±0.56 | 94.63 ±0.38** | 86.02 ±0.400** | 30.03 ±0.43** |
| miRNA-145a | 20.65 ±0.52 | 85.31 ±0.53** | 80.74 ±0.59** | 20.64 ±0.57 |
| miRNA-199a | 80.23 ±0.72 | 330.38 ±0.74** | 311.98 ±0.72** | 66.16 ±0.44a |
Correlation of the different serum microRNAs levels in HCC patients with AFP and DCP levels
The AFP and DCP are directly correlated with miR-139, miR-145, miR-199a, miR-122 and miR-125a (Table III). The results revealed a positive correlation between serum levels of miR-145 and miR-139 in HCC patients. Moreover, the serum levels of miR-199 were positively correlated with miR-139 and miR-145. However, a reverse correlation between miR-122 and miR-125a was also observed (Table IV).
Table III
Correlation of different serum microRNAs levels in HCC patients with AFP and DCP levels
ROC curve and CI analysis
Receiver operator characteristic curves (ROC) and confidence intervals (CI) of miR-122a, -125a, -139, -145 and -199a were established to discriminate HCV-induced HCC from CHC. MiR-122a (AUC = 0.897; 95% CI: 0.841–0.953), miR-199a (AUC = 0.889; 95% CI: 0.843–0.955), miR-125a (AUC = 0.780; 95% CI: 0.683–0.878), miR-139 (AUC = 0.930; 95% CI: 0.885–0.975) and miR-145 (AUC = 963; 95% CI: 0.932–0.994) (Figure 1).
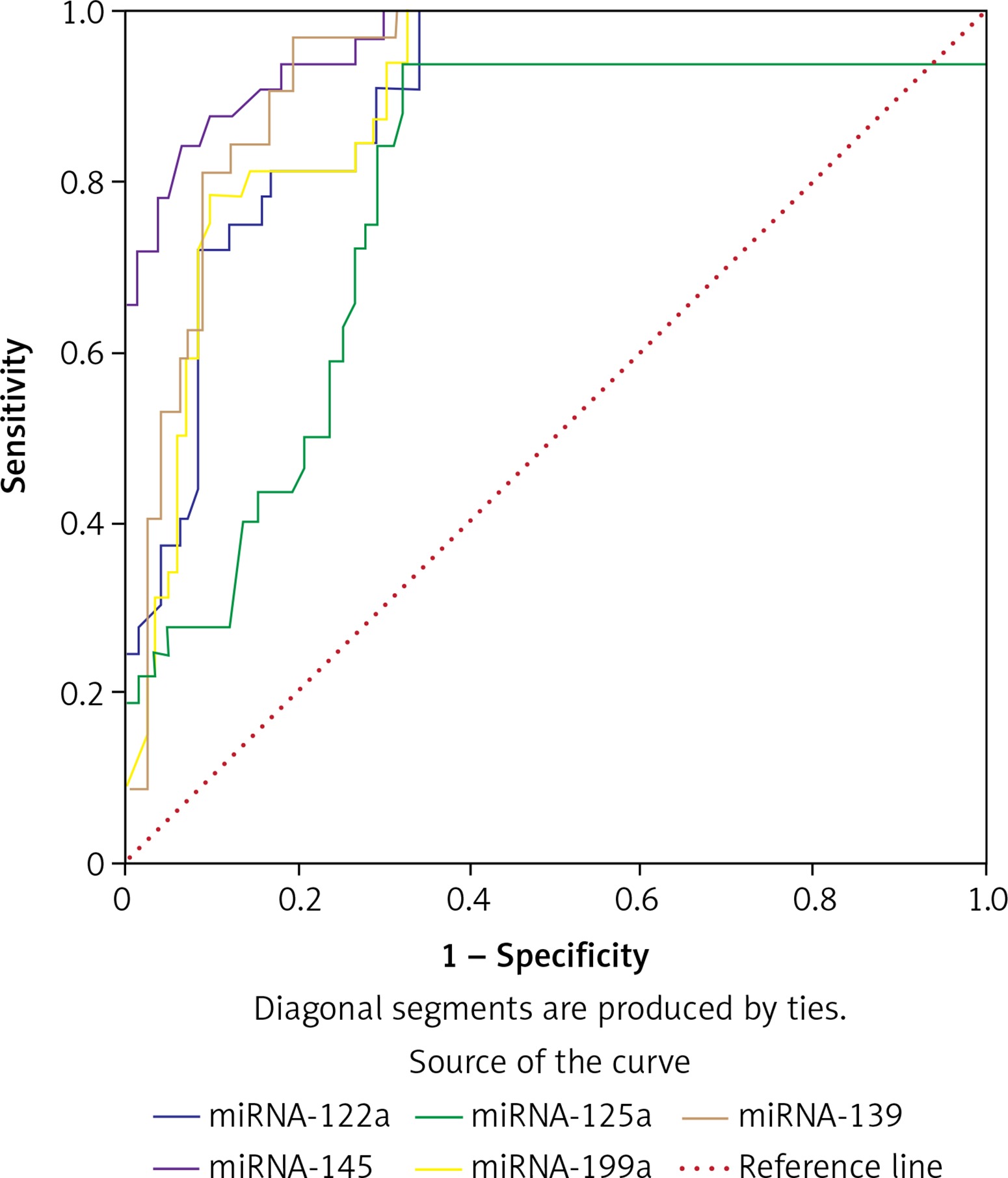
Diagnostic performance of circulating microRNAs in predicting HCC
To evaluate the usefulness of the studied serum biomarkers for predicting significant HCC, the area under the sensitivity, specificity, positive predictive value (PPV) and negative predictive value (NPV) were analyzed for each microRNA (Table V).
Discussion
Egypt has the highest prevalence of HCV infection in the world, estimated nationally as 14.7%, with genotype 4 being the most common [16]. Hepatocellular carcinoma is still an extremely poor prognostic cancer that remains one of the most widespread and aggressive human malignancies worldwide. Uncontrolled cell growth and neoangiogenesis are key features of occurrence, invasion, metastasis and malignant progression of HCC [17, 18]. The early diagnosis of HCC is of great clinical desirability since it promises good prognosis if the patients could get early surgical treatment. Until now, AFP has been mainly used clinically for the diagnosis of primary HCC; however, its sensitivity and specificity are not satisfactory [19]; therefore, novel biomarkers for early HCC diagnosis are gravely needed.
Results from recent studies revealed that circulating miRNAs are potential diagnostic biomarkers and prognostic factors in various kinds of diseases, especially in the field of human carcinogenesis. Chen et al. [20] demonstrated that by exploiting serum directly or by extracting RNA from the serum, it is possible to identify unique miRNA expression profiles specific for lung cancer, colorectal cancer and diabetes patients compared with healthy subjects.
In the past few years, miRNAs have been under the spotlight in the field of molecular oncology for their ability to be used as a powerful diagnostic tool.
Though several research groups have studied the role miRNAs play in the process of HCC development, the majority of the reports have studied histological specimens, therefore presenting limited clinical significance. Since miRNAs are shown to be stable in circulation, we propose that they can be utilized as a powerful non-invasive diagnostic tool for HCC. Therefore, we analyzed the expression profiles of five miRNAs (miRNA-122a, miRNA-125a, miRNA-139, miRNA-145, and miRNA-199a) in 165 individuals, and assessed their diagnostic ability compared to AFP and other conventional HCC markers. All patients had HCV genotype 4 since the viral genotype has an impact on miRNAs expression pattern as it has been previously described that many miRNAs were significantly different between CHC patients with HCV genotype 1 (GT-1) and non-genotype-1 (non-GT-1) [21].
The expression of miRNA-122a is specific to the liver, in which it is considered to represent 70% of the entire miRNA expression in the liver. Current studies have shown that the expression levels of miRNA-122a have declined during the process of hepatocarcinogenesis; hence miRNA-122a can function as a tumor suppressor. Our findings were consistent with previous studies, in which we observed a decline in the levels of miRNA-122a in HCC patients compared to the control individuals [22–24]. On the other hand, the levels of miRNA-122a increased significantly in the CHC and LC groups, suggesting that during the process of hepatocyte injury miRNA-122a levels increase drastically, and then decline back again significantly after the liver has entered carcinogenesis. According to a study by Jiang et al. [25], the serum levels of miRNA-122a have been shown to be elevated in HCC patients compared to healthy individuals. Also, as presented in our results, the ROC curve has demonstrated that out of the five miRNAs investigated, miRNA-122a has the highest specificity and sensitivity, making it an eligible candidate as a liver tumor marker. We inferred that the high AUC values are due to the fact that the patients in our studies had late stage HCC.
Halász et al. [26] have shown that in stage F4 fibrosis, the level of miR-122 was reduced as compared to stage F0, in which miR-122 showed a negative correlation with fibrosis stage in fibrotic liver samples and with LS values. These findings are supported by studies that discovered a negative correlation between miR-122 and fibrosis stage in chronic HCV infection, HCV-based HCC, and cirrhosis [27], in addition to the observations of a diminished level of miR-122 in non-alcoholic fatty liver disease (NAFLD) [28] and in HCC studies [29].
Loss of miR-122 appears to be a frequent event in HCC [30], which correlates with migration, invasion and in vivo tumorigenesis, whereas HCC cells expressing miR-122 maintain an epithelial phenotype [31]. MiR-122 is considered as a differentiation marker for hepatocytes [32], and a lower level of miR-122 might also reflect a lower degree of differentiation in the embryonal component. In addition, reduced expression of miR-122 is essential for the normal hepatocyte function; it positively regulates cholesterol, triglyceride accumulation, and fatty acid metabolism [33], constituting 70% of the total miRNA pool of the liver.
The majority of studies that focused on the expression profiles of miRNA in HCC have shown that in most cases the progression of the malignancy is correlated with the down-regulation of the miRNAs. However, it needs to be taken into consideration that at the post-transcriptional level, miRNAs regulate hundreds of targets that are part of many signal transduction pathways, which makes the role of miRNAs in the process of hepatocarcinogenesis very complicated. According to different studies, it has been shown that miR-125a is down-regulated in a number of tumors, such as medulloblastoma, breast cancer, lung cancer, gastric cancer, and ovarian cancer. Bi et al. [34] showed that the expression of miR-125a is lower in HCC malignant tissue compared to non-tumor adjacent liver tissue and that the low expression level of miR-125a was associated with the progression of the disease as well as poor prognosis. In addition to the lower expression observed in tissues, also low expression of miR-125a was seen in HCC cell lines. In fact, it has been shown that the in vitro and in vivo ectopic expression of miR-125a can lead to inhibition of proliferation, migration, and invasion of the HCC cells [34]. Unlike what the literature and other studies have shown, the expression level of miR-125 in our study was slightly higher in the HCC patient samples compared to the control group. On the other hand, very high expression levels of miR-125a were observed in the CHC and LC patient groups.
As with miR-125a, according to previous studies the expression level of miR-139 has been shown to be significantly lower in HCC patient samples compared to control individuals. MiR-139 was shown to play an important role in hepatocarcinogenesis. In fact, high expression levels of miR-139 were shown to inhibit cellular proliferation and invasion in malignant liver cells. In addition, in miR-139-transfected cells, apoptosis was shown to be highly induced, which suggests that miR-139 can act as a tumor suppressor through the inhibition of cell proliferation and invasion in HCC [35]. As seen in our results, the expression levels of miR-139 were higher in the HCC patient group compared to the control group, which contradicts what has been reported in other studies, while the CHC and LC patient groups showed a significant increase in miR-139 expression. As a matter of fact, a possible explanation for the elevated expression levels for miR-125a and miR-139 in the HCC patient group is that HCV infection is the underlying cause of the HCC cases in the study, which might have an effect on the expression level of the two miRNAs. Although according to the literature, HCV infection should not affect the expression levels after the patient develops HCC, no study has examined that with HCV genotype 4. All patients in our study have developed HCC as a result of a genotype 4 HCV infection, which may have resulted in the high expression level of miR-125a and miR-139, contradicting other studies.
MiR-145 was another miRNA tested among the miRNA panel investigated, in which its expression level showed no difference between the normal controls and the HCC patient group. As with the rest of the miRNAs investigated, miR-145 in the CHC and LC groups showed a significant increase compared to the control and HCC groups. According to Wang et al. [36], miR-145 was significantly down-regulated in HCC patient samples as well as HCC cell lines. As a matter of fact, the down-regulation of miR-145 was strongly linked to intrahepatic metastasis, tumor size, vascular invasion, and tumor grade, all suggesting that miR-145 acts as a tumor suppressor and its decreased expression results in progression of hepatocarcinogenesis.
The last miRNA we investigated was miR-199a, which was down-regulated in the HCC patient group compared to the control group, while it was significantly upregulated in the CHC and LC groups. The down regulation of miR-199a has been reported in several studies; in fact, miR-199a has been one of the most consistently reported miRNAs to be involved in HCC. MiR-199a is the third highly expressed miRNA in the liver, and in the case of HCC, it has been shown to be down-regulated especially in patients with HCV, HBV infections, and alcohol abuse [37]. The down-regulation of miR-199a in HCC patients has been associated with poor prognosis. The identification of miRNAs associated with HCC is crucial to developing new diagnostic and therapeutic tools with high specificity and sensitivity in order to combat this vicious human cancer.
In conclusion, our results indicate that measurement of serum levels of miRNA-122a, miRNA-125a, miRNA-139, miRNA-145 and miRNA-199a can help to differentiate HCC from CHC and LC. Also, they suggest that serum miR-122 might serve as a novel potential non-invasive biomarker for HCV-induced HCC.