Introduction
The incidence and mortality rates for the majority of cancers are decreasing in most western countries, but in less developed and economically unstable countries they are increasing [1]. Melanoma is a type of cancer which develops from melanocytes [2]. Melanomas generally occur in the skin and are rarely found in mouth, intestines and eye. Exposure to ultraviolet light (UV) is one of the main causes of melanoma, and the risk of UV-induced melanoma is higher people with low levels of skin pigmentation [2]. The UV light may be from either the sun or from other sources, such as tanning devices [3]. Genetic defects such as xeroderma pigmentosum also increase the risk of melanoma. Surgery, radiotherapy, chemotherapy, immunotherapy and biotherapy are the treatment regimens for melanoma [4]. Melanoma is one of the most dangerous types of skin cancers. Globally, in 2012, it was detected in 232,000 people and resulted in 55,000 deaths. Australia and New Zealand have the highest rates of melanoma in the world. There are also high rates in Europe and North America, while it is less common in Asia, Africa, and Latin America. Melanoma is more common in men than women [4]. When the whole genomes of 25 melanomas were sequenced, an average of about 80,000 mutated bases, mostly C > T transitions and about 100 structural rearrangements, were found in each melanoma genome. This is much higher than the approximately 70 mutations across generations from parent to child [5].
Flavonoids are ubiquitous polyphenolic compounds present in plants and include an important part of the human diet all through the course of evolution. More than 4,000 distinct, natural flavonoids have been recognized thus far and have been consumed as part of common foods, wines, teas, traditional medicines and pharmaceutical drugs [6]. They exhibit properties valuable for human health because they interact with several cellular targets. Flavonoids show anti-oxidant and free-radical scavenger activities along with anti-inflammatory, antiviral, and particularly anti-cancer properties [7]. Chemoprevention of cancer using either natural compounds or synthetic materials has now become a very important issue in the management of carcinogenesis. Wogonin is an O-methylated flavone, a flavonoid-like chemical compound which has been isolated from Scutellaria baicalensis. The glycosides of wogonin are known as wogonosides [8]. Preliminary in vitro studies have shown pharmacological effects indicating that wogonin may have anti-tumour properties [9, 10]. Wogonin has also been found to possess anticonvulsant effects. It acts as a positive allosteric modulator of the benzodiazepine site of the GABAA receptor [11]. The main purpose of the current research work was to evaluate the anticancer effects of methylwogonin in A375 human melanoma cells and to investigate its effects on apoptosis, DNA fragmentation, cell invasion and the PI3K/Akt signalling pathway.
Material and methods
Chemicals and other reagents
Methylwogonin (≥ 95%), MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide), and dimethyl sulfoxide were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Acridine orange, propidium iodide and Hoechst 33342 were procured from Wuhan Boster Biological Technology Ltd. (Wuhan, China). Dulbecco’s modified Eagle’s medium and RPMI-1640 medium were obtained from Gibco Life Technologies, Grand Island, NY, USA). Heat-inactivated fetal calf serum, penicillin, and streptomycin were obtained from Thomas Scientific, High Hill Road, Swedesboro, U.S.A.
Cell line and cell culture conditions
A375 human malignant melanoma cells were purchased from the cell bank of the Basic Medical College of Huazhong University of Science and Technology (HUST). The cells were maintained in DMEM supplemented with 10% FBS and antibiotics (100 U/ml penicillin G and 100 μg/ml streptomycin) at 37°C in a humidified incubator.
MTT assay for cell proliferation
The cell cytotoxicity induced by methylwogonin was estimated by MTT cell viability assay using different doses of the drug and at different time incubations. Briefly, A375 human malignant melanoma cells were plated at a density of 1 × 106 cells per well in 96-well plates for 12 h. The cells were then treated with 0, 5, 10, 25, 50, 150 and 300 μM methylwogonin for 24 and 48 h periods. MTT solution (20 μl) was added to each well. The medium was completely removed and 500 μl of DMSO was added to solubilize MTT formazan crystals. The optical density was determined at 570 nm (OD570) using an ELISA plate reader (Model 550; Bio-Rad, Hercules, CA, USA).
Clonogenic assay (colony forming assay) and determination of melanin content
A375 human malignant melanoma cells (2 × 106 cells/well) were plated into a 6-well plate for adherence for 12 h prior to drug treatment. After the cells had adhered, the cells were subjected to the treatment of different doses (0, 50, 150, 300 μM) of methylwogonin for 48 h. After this time interval, the used medium was discarded and the A375 cells were allowed to make colonies in complete medium for one week, after which colonies were fixed with acetic acid solution for 10 min, stained with Giemsa for 15 min and then the cells were counted manually under a light microscope. The melanin content of the melanoma cells was measured by the method as described previously by Hosoi et al. [12].
Fluorescence microscopic study of apoptosis using acridine orange/propidium iodide double staining
The apoptosis induced by methylwogonin in A375 human malignant melanoma cells was assessed by a fluorescence microscope using a double staining dye (acridine orange/propidium iodide). The A375 cells were seeded in 6-well plates at a density of 2 × 105 cells/well. The cells were treated with increasing concentrations of methylwogonin (0, 50, 150, 300 μM) for 48 h. Afterwards, the treated and untreated cells were incubated with acridine orange and propidium iodide (20 μg/ml each) for 1 h. The cell morphology was finally examined under a fluorescence microscope (Olympus IX 81, Tokyo, Japan) at 400× magnification.
Fluorescence microscopic study of apoptosis using Hoechst 33342
The A375 human malignant melanoma cells were treated with increasing concentrations (0, 50, 150, 300 μM) of methylwogonin and the cells were placed in a CO2 incubator for 48 h at 37°C. After incubation, the cells were fixed with 2.5% formaldehyde for 30 min and washed with PBS twice. The solution of Hoechst 33342 was added to the cells and after 20 min of staining the cells were observed under a fluorescence microscope at 200× magnification (Olympus IX 81, Tokyo, Japan).
DNA fragmentation analysis
In brief, A375 human malignant melanoma cells were seeded in a 60-mm cell culture plate, incubated for 48 h and then treated with 0, 50, 150, and 300 μM of methylwogonin for 48 h. Subsequently the A375 cells were harvested and washed twice with PBS before the pellets were lysed with a DNA lysis buffer for 30 min. The sample was centrifuged at 12,000 rpm and the supernatant was prepared in an equal volume of 3.5% sodium-dodecyl sulphate, then incubated with 10 mg/ml RNase A for 2 h. After adding 10 M ammonium acetate, the DNA was precipitated with ethanol and collected by centrifugation at 12,000 rpm for 30 min. Finally, the DNA was dissolved in gel loading buffer, separated by electrophoresis in 3.5% agarose gel, stained with ethidium bromide and examined under UV.
Invasion assay
After the A375 human malignant melanoma cells were washed with PBS, the cells were again suspended in serum free medium. Then 200 μl of cell suspension (2 × 106 cells/ml) was added to the upper chamber coated with Matrigel membrane (Millipore, Billerica, MA, USA) while the lower chamber was filled with medium containing 5% FBS. The cells were incubated with increasing concentrations (0, 50, 150, and 300 μM) of methylwogonin for 50 min at 25°C. After 48 h, the cells which still remained in the upper face of the filters were removed, and the cells which had travelled to the lower face of the filters were fixed with 90% methanol and then stained with 0.35% crystal violet and then counted using a light microscope.
Determination of melanin content
Western blot analysis
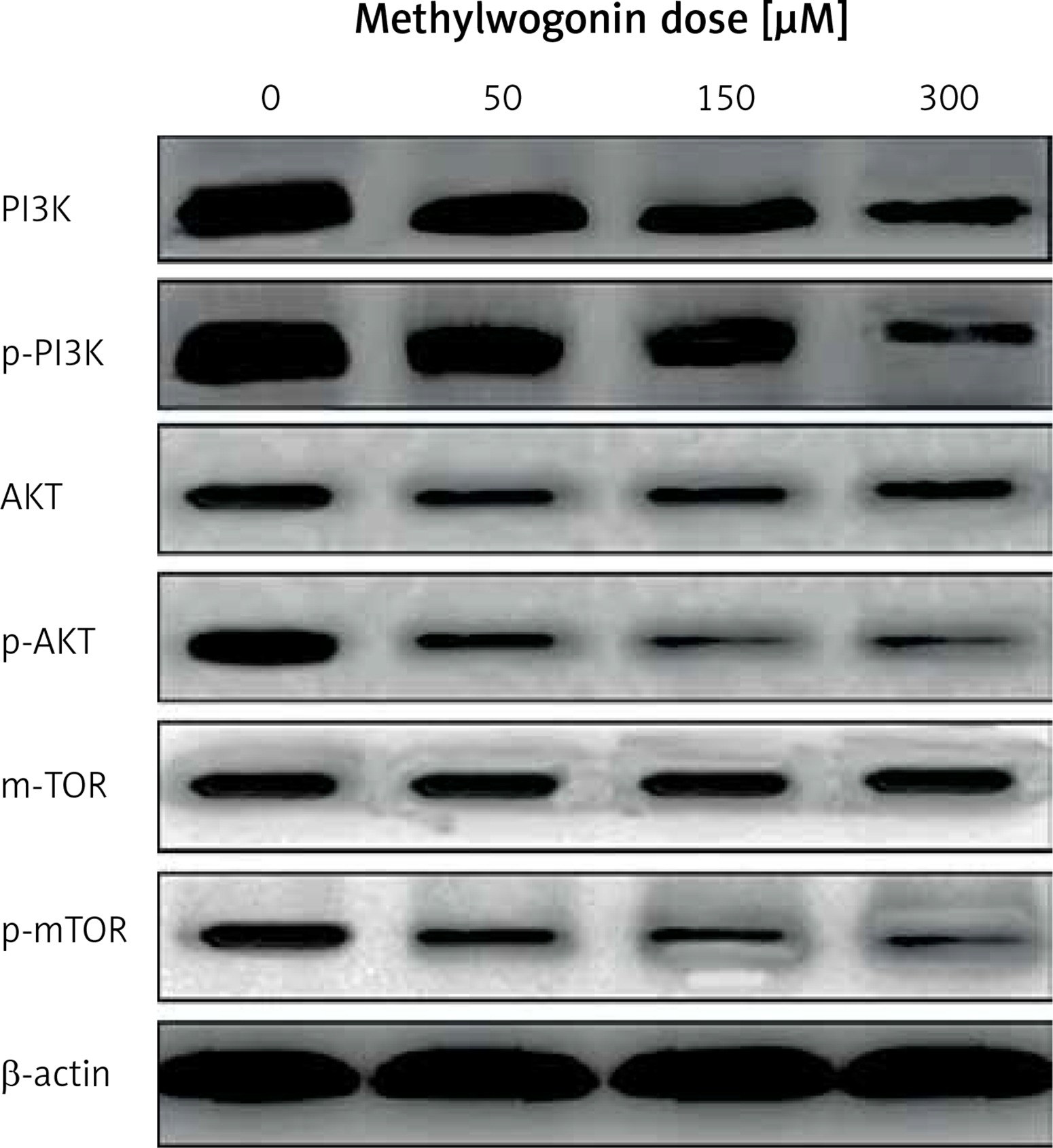
Next we used western blot assay to determine the main proteins involved in the anticancer effect produced by methylwogonin. A375 human malignant melanoma cells were harvested and lysed with RIPA buffer and the collected protein samples were examined using a bicinchoninic acid (BCA) protein assay kit for protein estimation. About 200 μg of cellular protein from each sample was applied to 8–10% SDS-polyacrylamide gels and probed with specific antibodies followed by exposure to horseradish peroxidase-conjugated goat anti-mouse antibodies. Blots were then developed using the West Pico Chemiluminescent substrate (Pierce; Woburn, MA, USA). The antibodies against β-actin, PI3K, Akt, p-PI3K, mTOR, p-mTOR, Bax, Bcl-2, and Cyt-c were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Statistical analysis
All results were depicted as mean ± standard error (S.E.M) from at least three independent experiments. The differences between groups were analysed by one-way ANOVA. Significance of differences was shown as *p < 0.05, **p < 0.01. The GraphPad prism 7 software was used for carrying out the statistical analysis.
Results
Methylwogonin induces potent cytotoxic effects in A375 human malignant melanoma cells
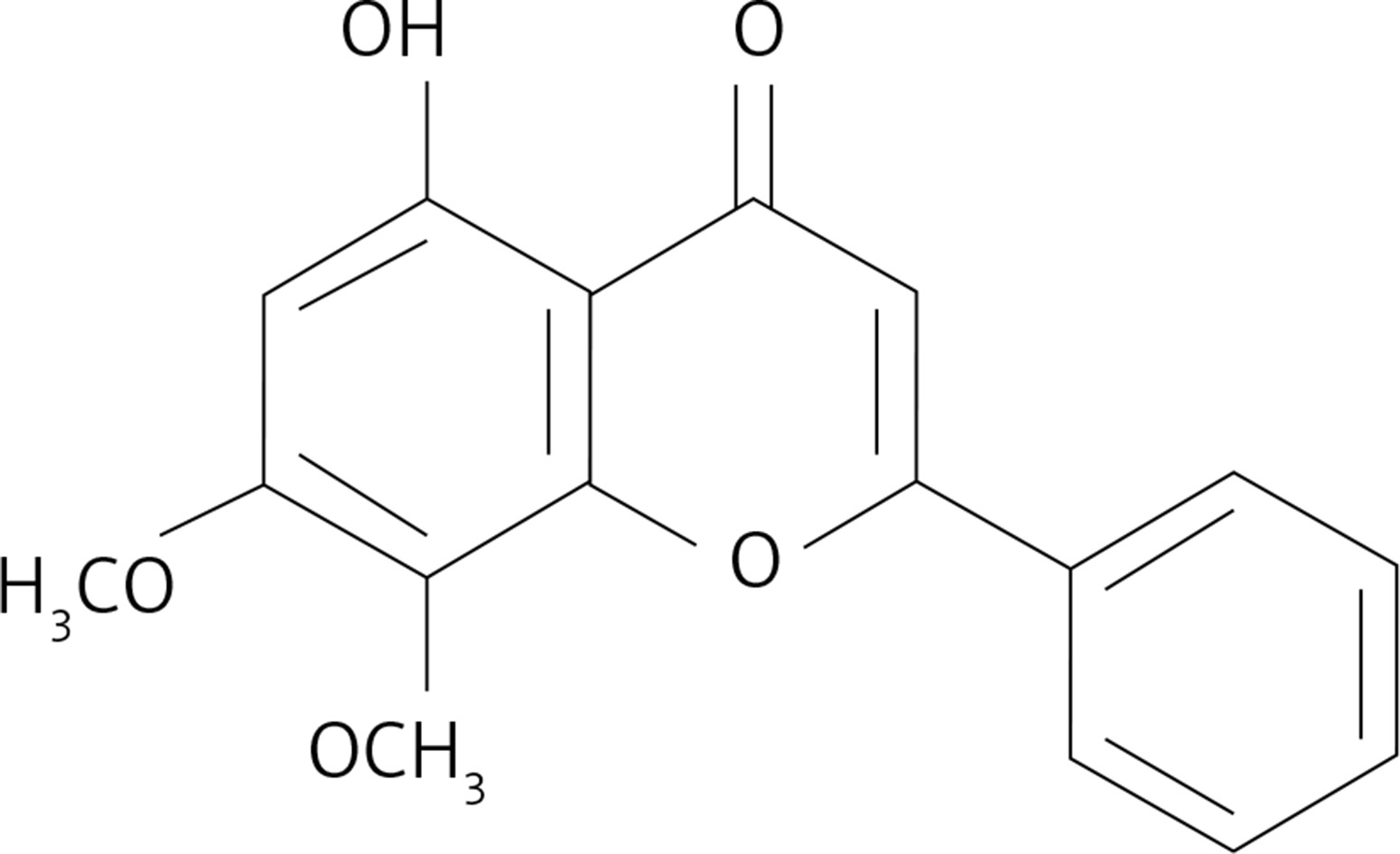
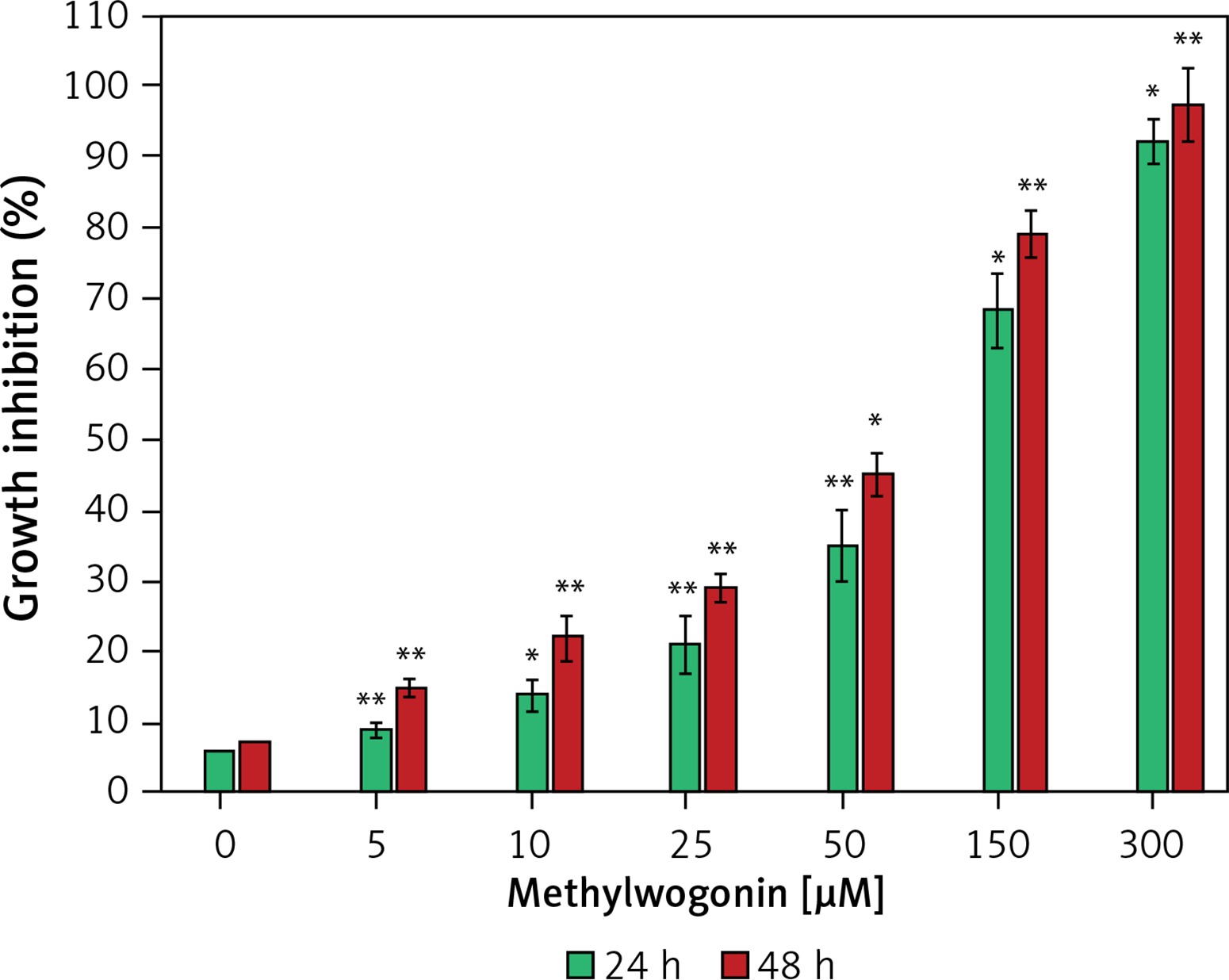
The fact that methylwogonin (Figure 1) induces strong cytotoxic effects in A375 human malignant melanoma cells was evaluated by MTT assay and the results are shown in Figure 2. As indicated in the figure, methylwogonin induces concentration-dependent as well as time-dependent growth inhibitory effects inducing significant cytotoxicity in these cancer cells. The IC50 values which were calculated at different incubation times were found to be 72.9 and 54.2 μM at 24 and 48 h, respectively. The half maximal inhibitory concentration (IC50) is a measure of the effectiveness of a substance in inhibiting a specific biological or biochemical function. Complete cell growth inhibition was observed at 48 h incubation with a 300 μM dose of the drug.
Methylwogonin led to inhibition of A375 cancer cell colony formation
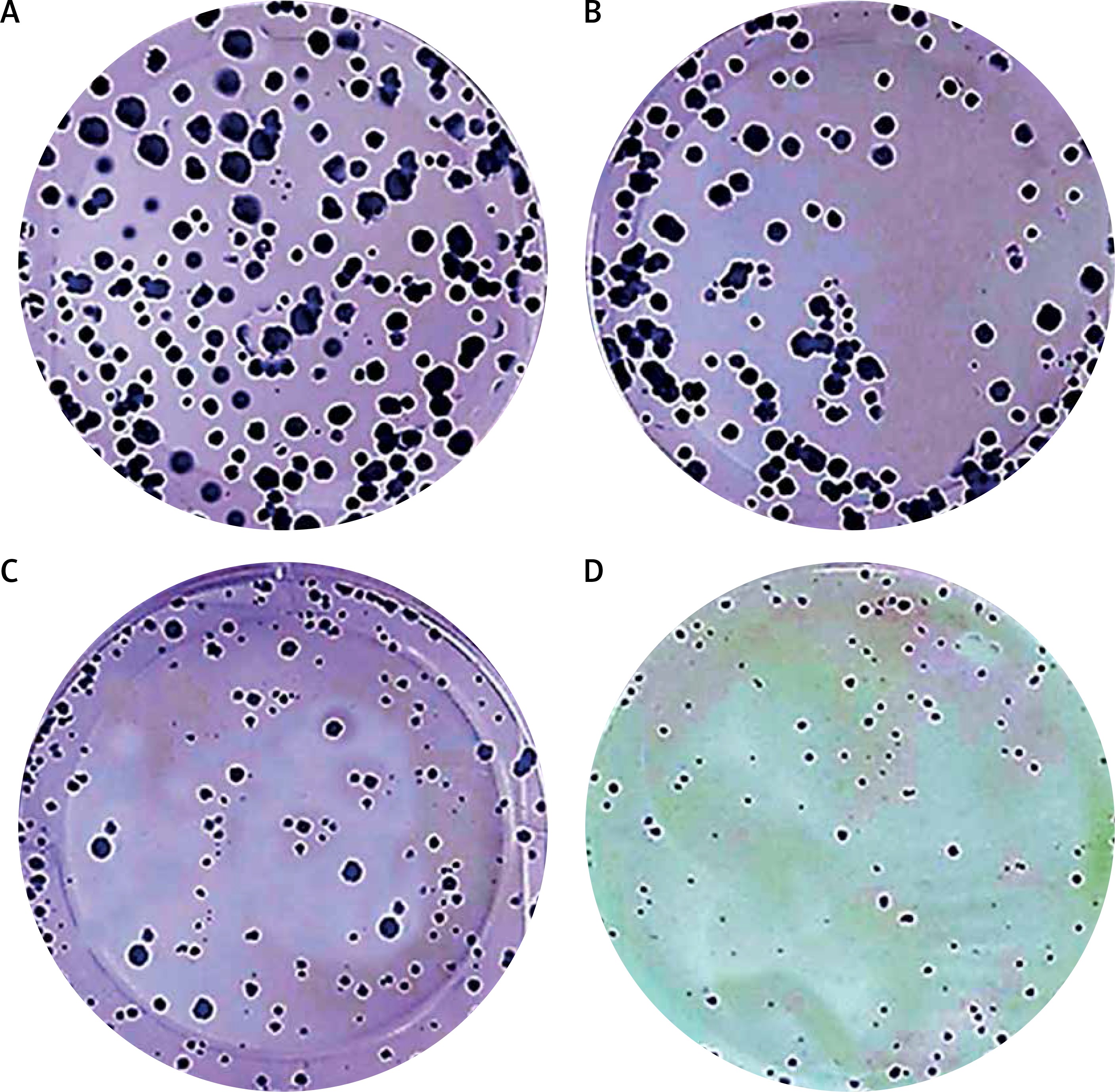
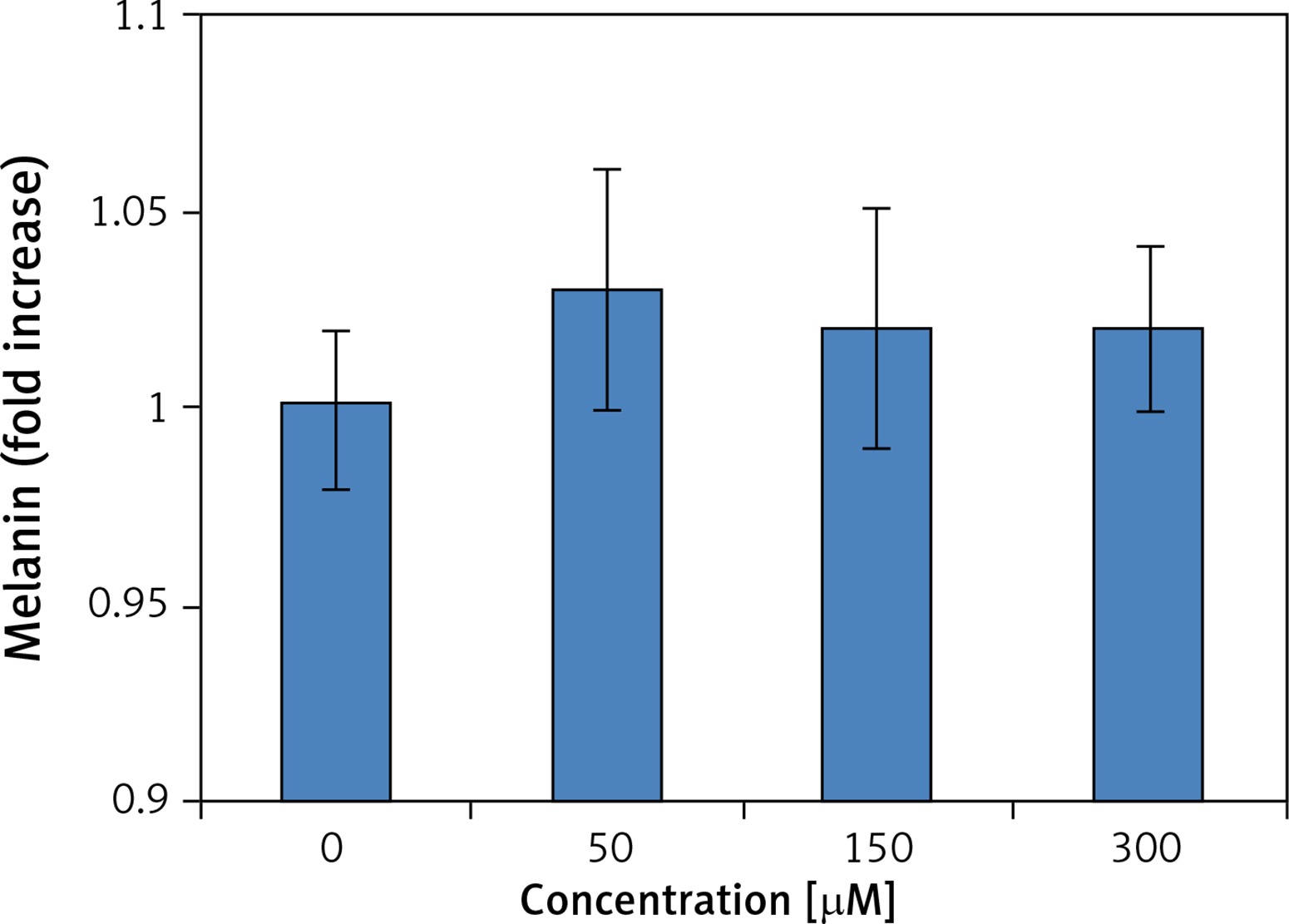
In order to evaluate whether methylwogonin induced both growth inhibitory effects, we used a clonogenic assay in order to assess effects of methylwogonin on the colony formation tendency in these cells. The results of this assay are depicted in Figure 3 and indicate that methylwogonin led to dose-dependent inhibition of colony formation in A375 human malignant melanoma cells. The number of cell colonies decreased significantly as the methylwogonin dose increased from 0, 50, 150, to 300 μM. The colonies decreased by 47, 64, 78% at the concentrations of 50, 150, 300 μM (p < 0.05) methylwogonin as compared to the untreated control. Thus taking both MTT and clonogenic assays into consideration, it is evident that methylwogonin induces cytotoxic effects along with inhibiting the colony formation tendency in these cells. Moreover, we also evaluated the melanin content of the cells, and no changes in melanin content were observed in the A375 cancer cells (Figure 4).
Methylwogonin led to apoptotic induction in A375 human melanoma cells
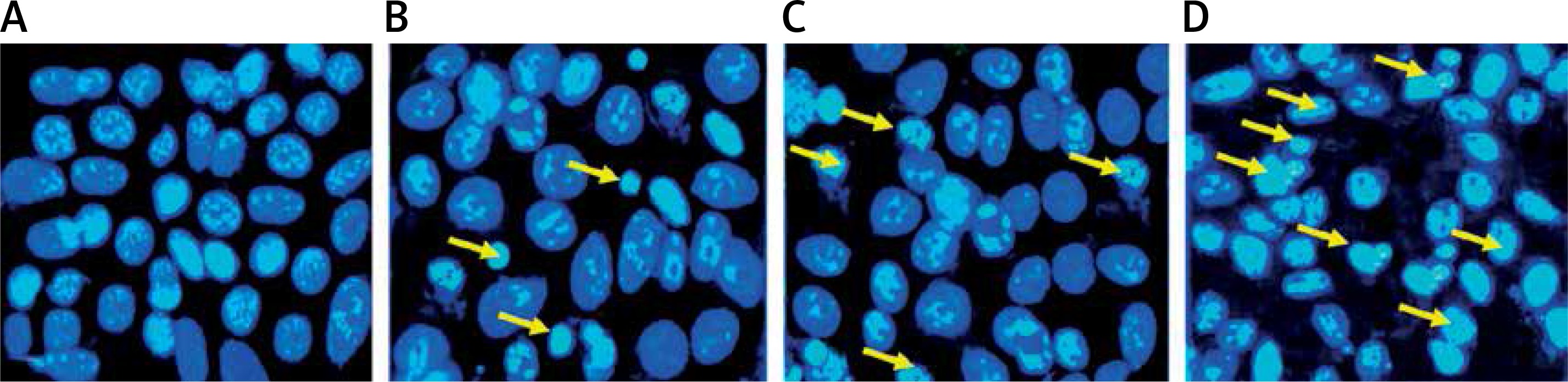
Further experiments were devised in order to evaluate the effects of methylwogonin on apoptotic induction in these cells using fluorescence microscopy with acridine orange/propidium iodide and Hoechst 33342 staining agents. The results which are shown in Figures 5 A–D and 6 A–D indicate that the untreated control cells showed prominent green fluorescence, indicating their viable nature (Figure 5 A). However, methylwogonin treatment of cells at lower doses led to yellow fluorescence (early apoptosis) which changed to red/orange fluorescence at higher doses, indicating late apoptosis (Figures 5 B–D). Lower doses of methylwogonin induced early apoptosis while higher doses of the compound led to late apoptosis. The apoptotic cell populations increased from 2.5% in the untreated control up to 85% at 300 μM (p < 0.01) concentration of methylwogonin. Similar results were obtained using Hoechst 33342 staining, and the results are depicted in Figure 6, indicating that untreated control cells showed normal cell morphology with no cell shrinkage. However, on treating cells with 50, 150 and 300 μM doses of methylwogonin, several morphological changes in the cells including chromatin condensation, fragmented nuclei and cellular shrinkage occurred. Methylwogonin-treated cells exhibited bright fluorescence indicating rupture of the cell membrane and extensive staining (Figures 6 B–D).
Figure 5
Fluorescence microscopic (400× magnification) study of the apoptosis induced by methylwogonin in A375 human malignant melanoma cells. The cells were treated with 0 (A), 50 (B), 150 (C), and 300 μM (D) doses of methylwogonin for 48 h and then stained with acridine orange/propidium iodide double dye and finally examined by fluorescence microscope at 400× magnification. White arrows indicate viable cells, orange arrows indicate early apoptotic cells, red arrows indicate late apoptotic cells
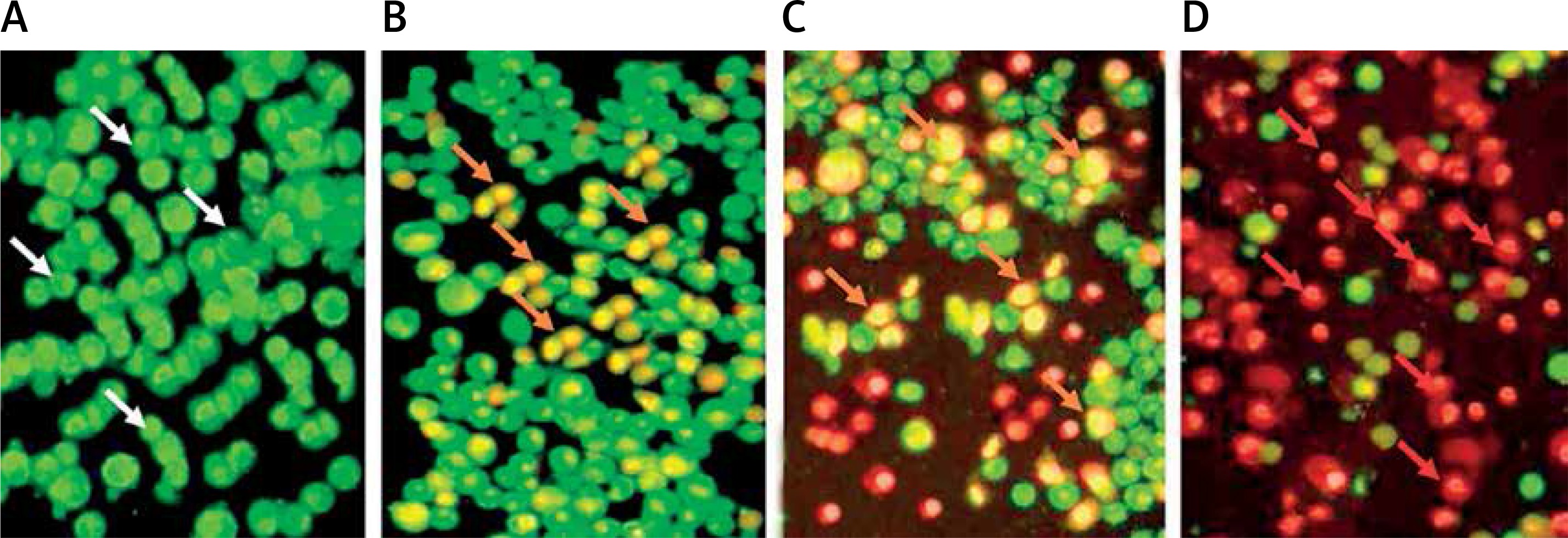
Figure 6
Fluorescence microscopic (at 200× magnification) study of apoptosis in A375 human malignant melanoma cells using Hoechst 33342 staining dye. The cells were treated with 0 (A, untreated cells), 50 (B), 150 (C) and 300 (D) μM concentrations of methylwogonin for 48 h and then stained with Hoechst 33342 before being analysed by fluorescence microscopy. Yellow arrows indicate cells with chromatin condensation, DNA fragmentation and cell shrinkage (characteristic of apoptosis)
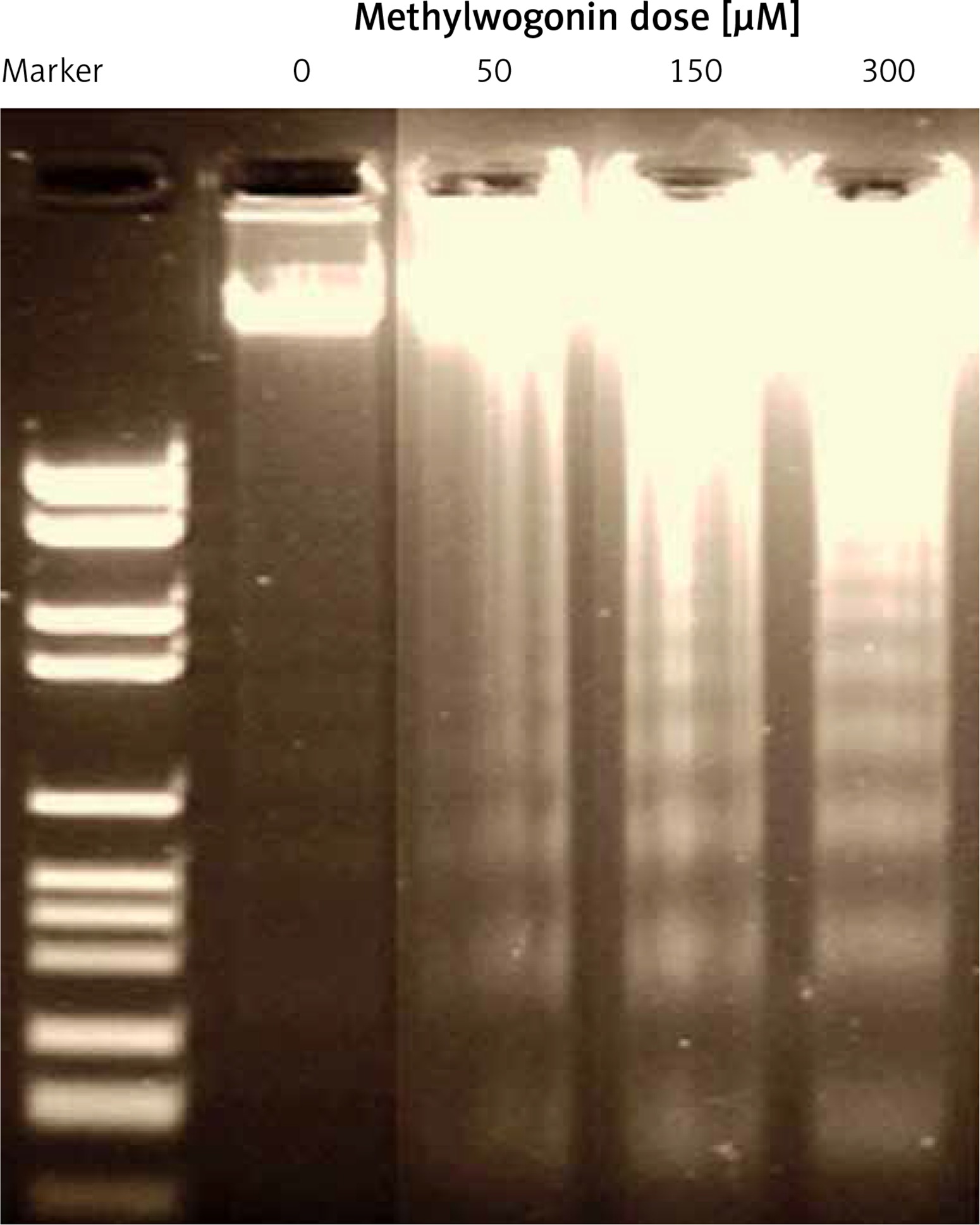
Methylwogonin led to DNA damage in A375 human melanoma cells
DNA damage which was induced by methylwogonin in A375 human melanoma cells was examined using gel electrophoresis, which involves the formation of a DNA ladder. The findings of this assay are depicted in Figure 7 and indicate that DNA ladder formation was observed and this effect increased with increasing doses of methylwogonin. In the case of the untreated control group, no such DNA laddering was seen, indicating no DNA damage at all. The DNA fragmentation is believed to be a hallmark of the apoptotic process which initiates within the cell, further indicating that methylwogonin led to cell death through inducing apoptosis.
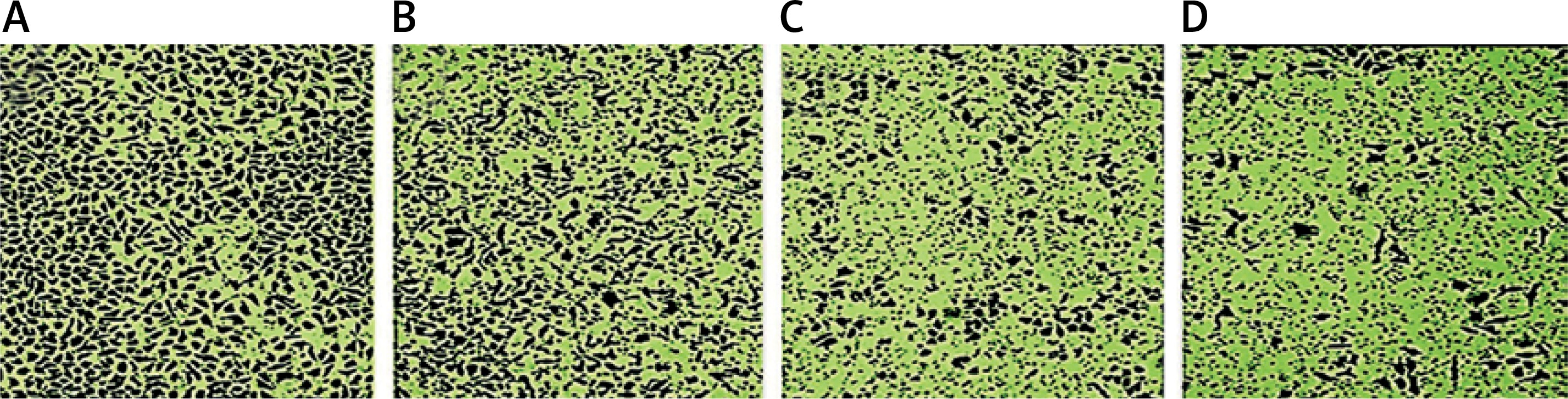
Methylwogonin suppressed cell invasion in A375 human malignant melanoma cells
The effect of methylwogonin on the cancer cell invasion in A375 human malignant melanoma cells was demonstrated by an invasion assay. The results, which are depicted in Figure 8, indicate that methylwogonin also inhibited cancer cell invasion in a dose-dependent manner. The percent inhibition of cell invasion was reported to be up to 69% at 300 μM (p < 0.01) concentration. Chemotherapeutic agents which suppress cancer cell migration and invasion are believed to be promising antitumor drugs because cancer cell migration and invasion have a direct relationship with cancer metastases.
Methylwogonin led to significant alteration of mTOR/PI3K/Akt signalling pathway
It is well established that the PI3K/Akt pathway has a close relationship with cell proliferation and cell survival. Therefore, our further aim was to evaluate the effects of methylwogonin on this crucial biochemical pathway. The results are depicted in Figure 9, which reveal that treatment of A375 human melanoma cells with different doses of methylwogonin led to concentration-dependent downregulation of phosphorylated PI3K, AKT and mTOR. However, it was found that total protein levels of PI3K, AKT and mTOR were not affected so much. Thus it now becomes evident that methylwogonin inhibited malignant melanoma cancer cell growth by regulating the mTOR/PI3K/AKT signalling pathway.
Discussion
Cancer is the second leading cause of death in Europe and America after cardiovascular disease. Discovery and development of anticancer drugs are the main emphasis of several pharmaceutical companies as well as non-profit government and non-government organizations. For many decades now, identification of cytotoxic compounds has led to the development of anticancer treatments. However, due to the development of multidrug resistance and emergence of severe side-effects, the use of cytotoxic anticancer drugs has been linked with limited efficacy, especially at advanced stages of the disease [11, 12]. There is an urgent need of such compounds which can selectively target cancer cells without causing too much damage to the normal cells. An ideal drug for the treatment of cancer is one which causes less harm to the normal cells. Induction of apoptosis may be useful in the therapy and management or prevention of cancer. Certain naturally occurring molecules induce apoptosis in cancerous cells but not in normal cells. Increasingly it has become obvious that apoptosis for many anti-tumour agents is an important mode of action [13–15].
Wogonin is an O-methylated flavone, which resembles flavonoids. It is extracted from S. baicalensis. Several previously published reports indicate that wogonin induces cytotoxic effects in various cancer cells including breast, osteosarcoma and malignant T cells via the induction of apoptosis and cell cycle arrest. It was also reported that up to a dose as high as 100 μM there was no cytotoxicity towards normal cells, indicating that wogonin induces selective cytotoxicity in cancer cells [9, 16–18]. It has also been reported that wogonin induced apoptosis via reactive oxygen species (ROS) generation along with modulation of PI3K/Akt, Bax and Blc-2-related signalling pathways. Studies also indicated that wogonin induced cytotoxicity in human promyeloid leukaemic cells. When tested it was found that wogonin was neither genotoxic nor apoptogenic in mice at indicated doses [19, 20]. Wogonin also led to induction of human glioma cell death, which was mediated by ROS generation, GPR78 and GRP94 expression, and increased activity of caspases, such as caspase-9 and caspase-3, eventually leading to apoptosis. Wogonin also inhibited cell mobility and invasion by upregulating the metastasis suppressor maspin gene [21, 22]. In summary, these data provide novel insights into the chemoprotective effect of wogonin, the main active ingredient of the Chinese medicine Scutellaria baicalensis. To the best of our knowledge, there are no reports on the antitumour effects of methylwogonin, which is a methylated derivative of wogonin. Previous studies have reported that melanin possesses radioprotective and scavenging properties and its existence can affect the susceptibility to the therapy, as shown in in vitro and in vivo experiments [23–26]. Therefore, we evaluated the effect of the test compound on the melanin status of the A375 human malignant melanoma cells (Figure 4). The results indicated the melanin content of the cells remained uniform throughout the experiment in all tested cell samples, and therefore it may be inferred that melanin content had no significant effect on the anticancer activity of methylwogonin the A375 human malignant melanoma cells. Further, we also studied the antitumor effects of methylwogonin on apoptosis induction, DNA damage, cancer cell invasion and the mTOR/PI3K/Akt signalling pathway. The MTT assay indicated that methylwogonin induced concentration-dependent as well as time-dependent growth inhibitory cytotoxic effects in these cancer cells. Moreover, the results of the MTT assay were also complemented by the results of the colony formation assay as the number of cell colonies decreased significantly with the increase in methylwogonin dose from 0, 50, 150, to 300 μM. Lower doses of methylwogonin induced early apoptosis while higher doses of the compound led to late apoptosis. Methylwogonin-treated cells showed evident signs of morphological changes characteristic of apoptosis including chromatin condensation, fragmented nuclei and cellular shrinkage. DNA ladder formation was observed on methylwogonin treatment and this effect increased with increasing doses of methylwogonin. Methylwogonin also inhibited cancer cell invasion in a dose-dependent manner. Previous studies have reported that the activation of the PI3K/AKT/mTOR pathway plays a vital role in ovarian cancer tumorigenesis, progression and chemotherapy resistance [27, 28]. Therefore we evaluated the effect of methylwogonin on this pathway. Treatment of A375 human melanoma cells with different doses of methylwogonin led to dose-dependent downregulation of phosphorylated PI3K, AKT and mTOR. However, it was found that total protein levels of PI3K, AKT and mTOR were not affected so much.
In conclusion, the present study revealed that methylwogonin induced potent antitumor effects in A375 human malignant melanoma cells by inducing apoptosis, DNA damage, inhibition of cell invasion and downregulation of the mTOR/PI3K/Akt signalling pathway.