Introduction
Diabetic nephropathy is one of the most common diabetic microvascular complications and a leading cause of end-stage renal disease [1]. There are many factors contributing to the development of diabetic nephropathy, but the major reason is the glomerular lesion [2–4]. Glomerular mesangial cells, which are one of the main parts of the renal glomerulus, are prone to hyperglycemia-induced cell apoptosis and injury [5–7]. In addition, loss of glomerular mesangial cells was correlated with the progression of diabetic nephropathy [8, 9]. Thus, mesangial cell apoptosis and injury play a crucial role in the progression of diabetic nephropathy. Hyperglycemia leads to mesangial cell apoptosis, possibly by triggering oxidative stress and the caspase-dependent mechanism of cell death [5, 6].
Published studies have shown that blood glucose fluctuation was associated with diabetic nephropathy in type 2 diabetes patients [10, 11]. A previous study reported that oscillating high glucose enhanced human coronary artery endothelial cell apoptosis [12]. Furthermore, in an in vitro study, blood glucose fluctuation exacerbated glomerular mesangial cells’ abnormal growth and collagen synthesis [13]. However, whether blood glucose fluctuation could contribute to mesangial cell apoptosis remains unclear.
As is known to all, the AKT network has an important effect on kidney cell apoptosis and cell injury [14, 15]. Phosphorylation of AKT protects mesangial cells against high glucose stimulation through inhibiting its downstream proteins, such as P-GSK-3β and cleaved caspase-3, and so on [16–19]. However, whether blood glucose fluctuation increases mesangial cell apoptosis related to inhibiting the AKT signaling pathway is still uncertain.
In the present study, we investigated whether glucose fluctuation could aggravate apoptosis in glomerular mesangial cells compared with sustaining high glucose stimulation, and its molecular mechanism.
Material and methods
Material
CCK8 was purchased from Dojindo (Kumamoto, Japan). Malondialdehyde (MDA) and superoxide dismutase (SOD) were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) to evaluate oxidative stress. 2′,7′-dichlorofluorescin diacetate (DCFH-DA), AKT, GSK3, caspase-3 and actin antibodies were obtained from Beyotime Institute of Biotechnology (Shanghai, China). P-AKT and P-GSK3β antibodies were purchased from Cell Signaling Technology, Inc (Shanghai, China). Dulbecco’s modified Eagle’s medium (DMEM/High Glucose) was obtained from Thermo Fisher Scientific (Shanghai, China) as well as DMEM/Low Glucose.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Cells cultures
Rat glomerular mesangial cells were bought from Boster (Wuhan, China). We selected the 5th–10th generation of mesangial cells in the present study. The cells were cultured in DMEM (5.6 mmol/l glucose) supplemented with 10% fetal bovine serum (FBS), antibiotics (100 U/ml penicillin and 100 mg/ml streptomycin), at 37°C in 5% CO2 for 24 h.
The cultured cells were randomly divided into four groups: the normal glucose group (NG, 5.6 mmol/l glucose), the high glucose group (HG, 25 mmol/l glucose, and then normal glucose conditions incubated for the night), the glucose fluctuation group (FG, alternated 5.6 mmol/l glucose and 25 mmol/l glucose every 3 h, repeated 3 times, and then normal glucose conditions incubated for the night) and the mannitol group (MG, 5.6 mM glucose plus 24.4 mM mannitol as an osmotic pressure control).
Effects of fluctuation of blood glucose on mesangial cell viability
Cell viability was assayed by the CCK8 kit according to the manufacturer’s protocol [20]. Briefly, mesangial cells were seeded onto 96-well plates and every well contained 5 × 103 cells. Then they were grown to 90% confluency. The cells were cultured for 12, 24, 48 and 72 h. Then, we added 10 µl of CCK8 reagent into every well. After 1–4 h of incubation at 37°C, cell viability was assessed by scanning with a microplate reader at 450 nm. There were at least 6 duplicate wells in each group.
Mesangial cell apoptosis using flow cytometry
Mesangial cell apoptosis can be detected using flow cytometry analysis [20]. In brief, the mesangial cells were seeded onto 96-well plates at 37°C in a humidified incubator containing 5% CO2 for 48 h. The cells were washed once in phosphate-buffered saline (PBS), collected by trypsin digestion with 0.25% trypsin (without EDTA), washed twice in PBS, then 500 µl of binding buffer, 5 µl of annexin V-FITC and 5 µl of propidium iodide were added to each well. Then, the cells were incubated in the dark for 15 min at 37°C and apoptosis was immediately analyzed.
Effects of fluctuating blood glucose on ROS levels in mesangial cells
The level of intracellular ROS was determined by flow cytometry using the peroxide-sensitive fluorescent probe 2′,7′-dichlorofluorescin diacetate (DCFH-DA) as previously described [21]. In brief, mesangial cells were seeded in a 96-well culture plates and incubated with 10 µM DCFH-DA for 30 min at 37°C. Then, cells were harvested and washed three times with PBS, re-suspended in ice-cold PBS and placed on ice in a dark environment, and washed twice with DMEM deprived of serum to fully remove unbound ROS from the intracellular DCFH-DA. The fluorescent signal was recorded using fluorescence microscopy (488 nm filter; OLYMPUS IX-73, Japan).
Effects of fluctuating blood glucose on MDA level and SOD activity in mesangial cells
The MDA level and SOD activity were analyzed using specific reagents according to the protocols provided by the manufacturer. Briefly, mesangial cells were seeded into 96-well plates and every well contained 5 × 103 cells. Cultured cells were incubated for 48 h. Then, we collected the supernatant to determine the MDA level and SOD activity.
Western blotting
Glomerular mesangial cells were plated in 100 mm dishes for western blot analysis. After 48 h cells were lysed with cell lysis solution. Lysate was centrifuged at 14 000 g for 30 min at 4°C, and the supernatant was stored at –80°C. The protein was loaded per lane, separated by SDS–PAGE gel and transferred onto an NC membrane and incubated with the primary antibody (1 : 500). After washing, the membranes were incubated for 2 h with secondary antibody (1 : 200). The immunoblots were scanned using Image-Pro Plus 6.0 (Media Cybernetics, Silver Spring, MD, USA).
Statistical analysis
At least six samples were selected from each group for experimental statistical analysis. Image J analysis software was used to analyze the Western blotting results. Data were expressed as mean ± SD. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by the least significant difference (LSD) test using SPSS 13.0 computer software. P < 0.05 was considered statistically significant.
Results
Mesangial cell viability under glucose fluctuation at different time points
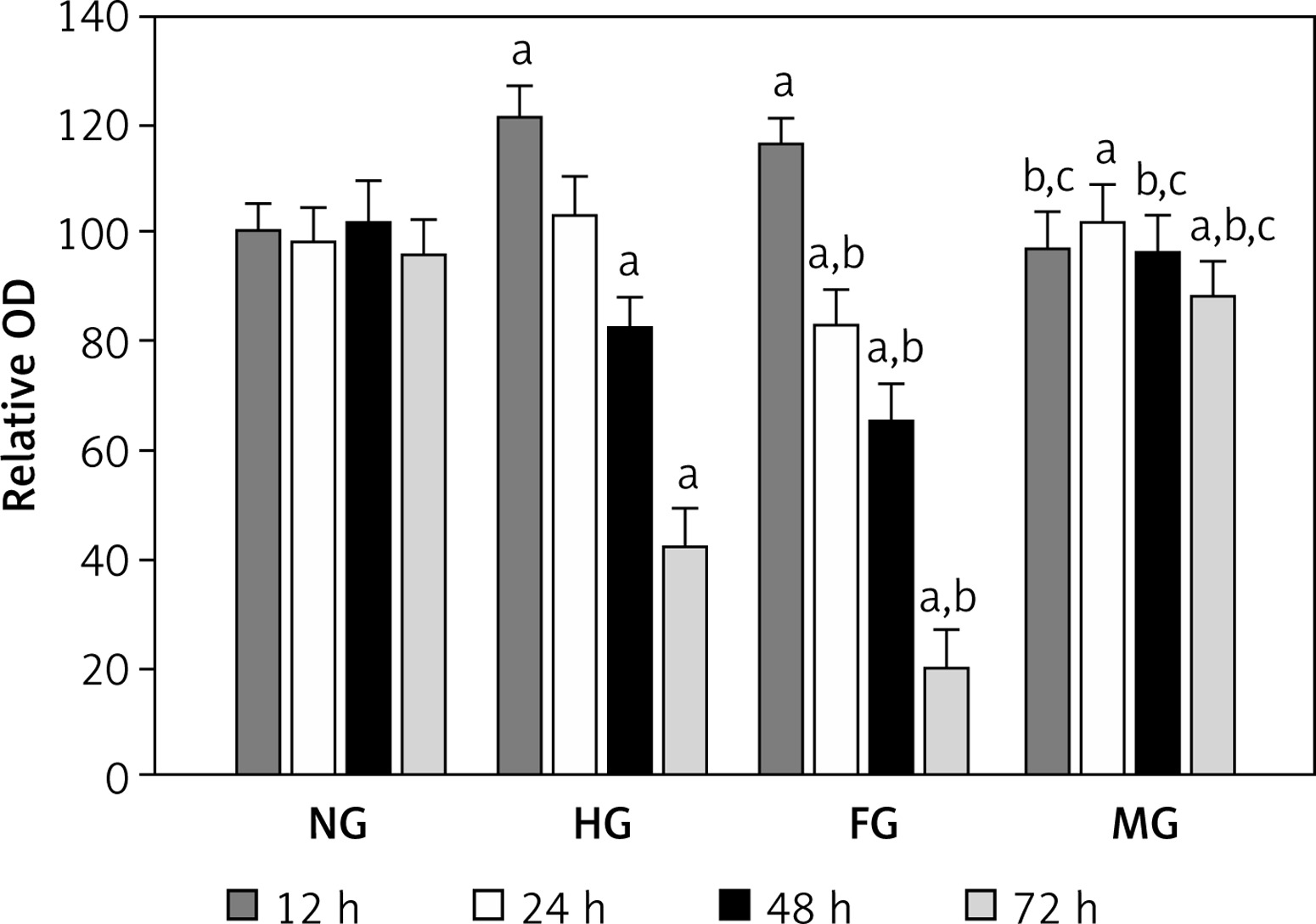
In order to elucidate the change of glomerular mesangial cells under glucose fluctuation at different time points, we examined glomerular mesangial cell viability using the CCK-8 method. As shown in Figure 1, mesangial cell viability in both the HG and FG groups was increased significantly in comparison with the NG group at 12 h, but there was no significant difference between the HG group and the FG group. However, mesangial cell viability was significantly decreased in the FG group compared with the NG and HG groups at 24 h, while no significant difference was found between the NG group and the HG group. When compared with the NG group, mesangial cell viability was decreased in the HG and FG groups at 48 h and 72 h; especially, the FG group had lower cell viability than the NG group. Moreover, there was no significant difference in mesangial cell viability between the NG group and the MG group at 12, 24 and 48 h. Nevertheless, mesangial cell viability in the MG group at 72 h was decreased significantly compared to the NG group. In general, these results indicated that fluctuating blood glucose obviously reduced mesangial cell viability from 48 h when compared with the HG group.
Figure 1
Effects of fluctuation blood glucose on mesangial cell proliferation. Cells incubated in different glucose conditions for 12, 24, 48 and 72 h were analyzed by CCK-8
Data are shown as mean ± SD. aP < 0.05 vs. the NG group, bp < 0.05 vs. the HG group and cp < 0.05 vs. the FG group by one-way ANOVA.
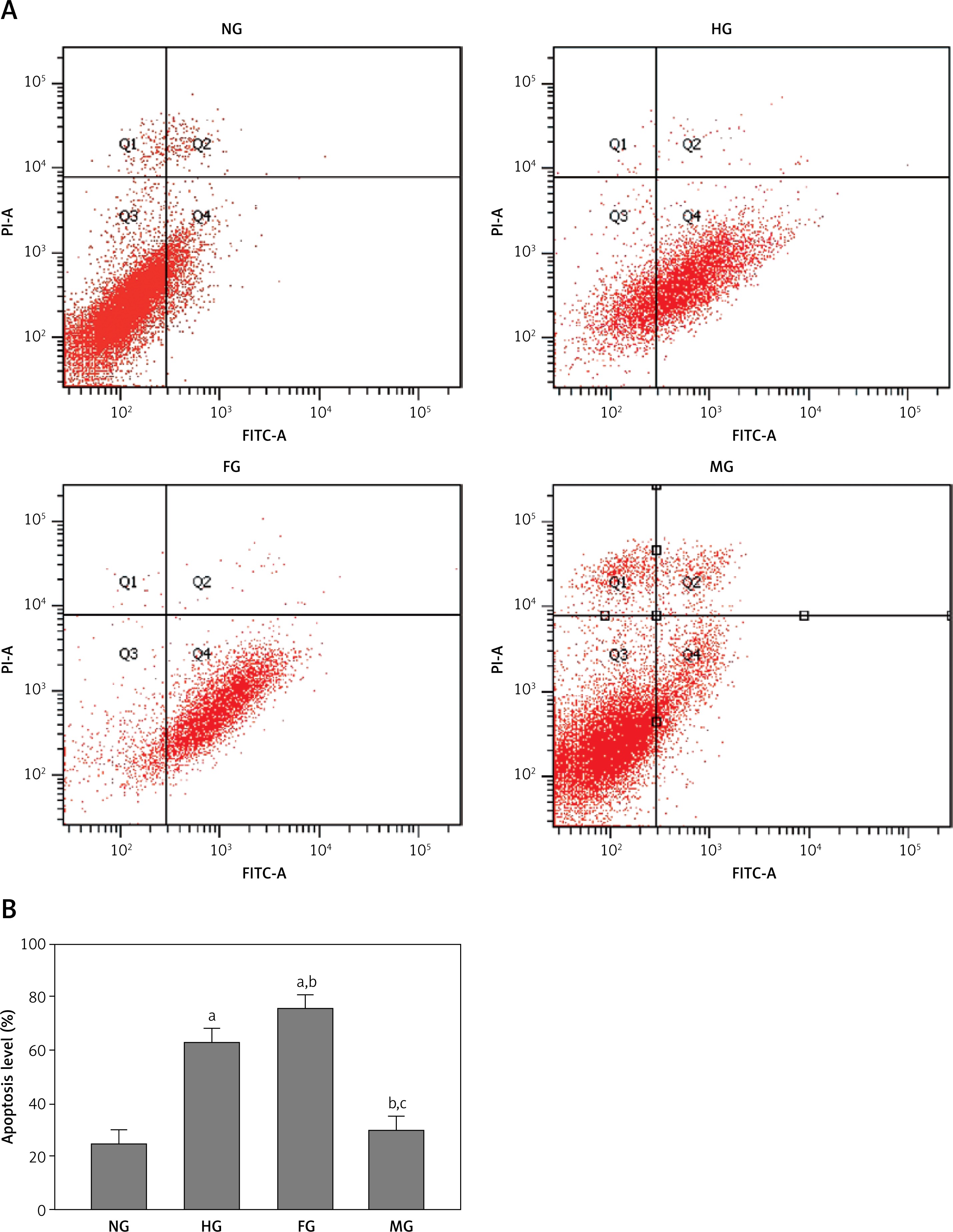
Fluctuating blood glucose increased glomerular mesangial cell apoptosis
Next, we detected glomerular mesangial cell apoptosis using flow cytometry. As shown in Figure 2, glomerular mesangial cell apoptosis in the FG group was significantly increased compared with either the NG group or the HG group, even though in the HG group it was higher than in the NG group. There was no significant difference between the NG group and the MG group. These results indicated that blood glucose fluctuation accelerated glomerular mesangial cell apoptosis.
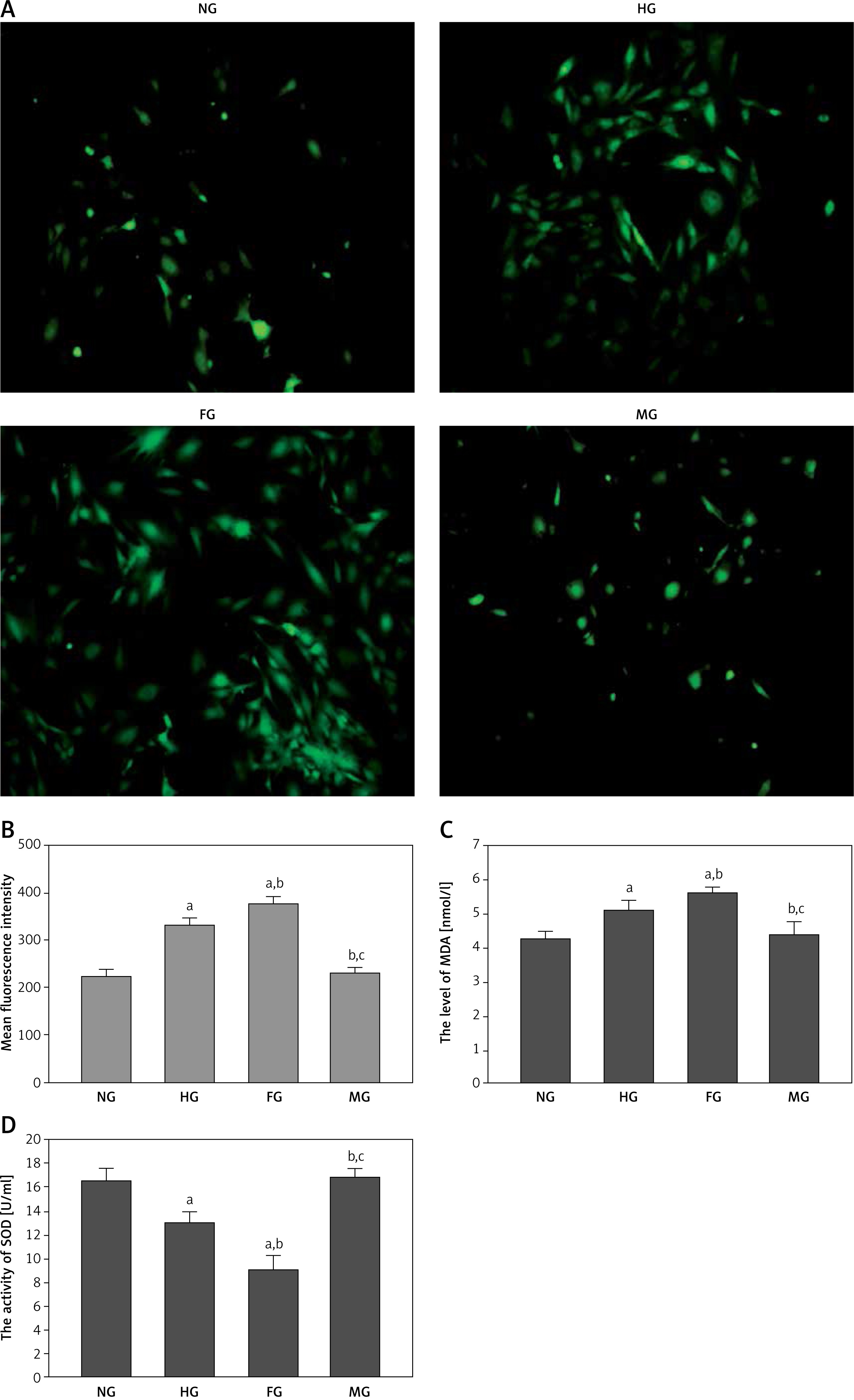
Glucose fluctuation aggravated oxidative stress in glomerular mesangial cells
It is reported that oxidative stress plays an important role in glomerular mesangial cell injury in high sugar conditions [22]. In addition, fluctuating blood glucose aggravated renal injury by increasing the generation of oxidative stress [23]. As shown in Figure 3, the level of intracellular ROS in the HG group was increased in comparison with that in the NG group; the increase was more obvious in the FG group, and the ROS level in the FG group was higher than in the HG group. The MDA level in the HG group was significantly raised compared with the NG group; moreover the increase was more marked in the FG group. The SOD activity was significantly lower in the FG group compared with either the NG group or the HG group. These results suggested that glucose fluctuation aggravated the oxidative stress state in glomerular mesangial cells.
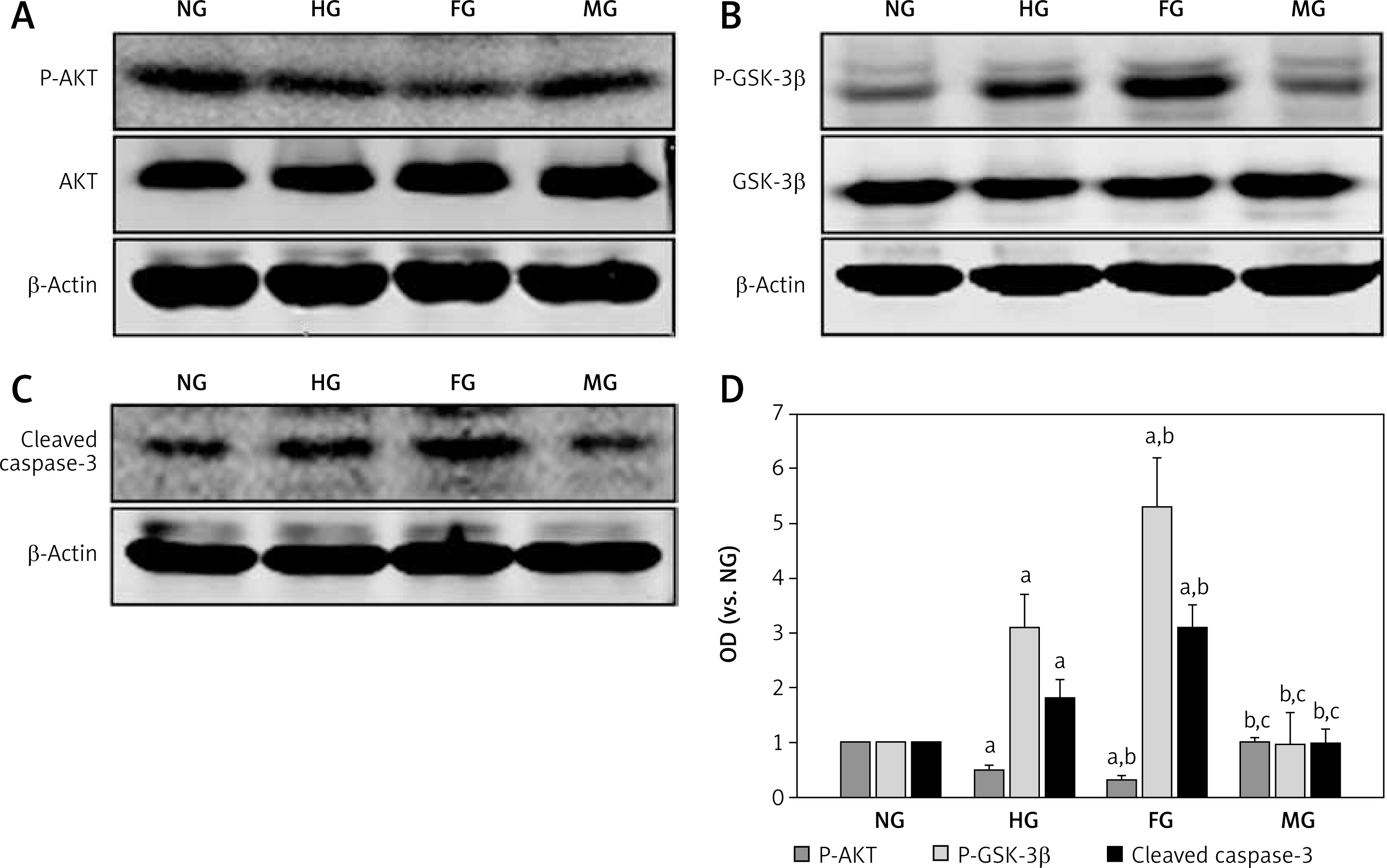
The effect of glucose fluctuation on P-AKT, P-GSK-3β and cleaved caspase-3 expression in glomerular mesangial cells
Previous studies indicated that high glucose accelerated the glomerular mesangial cell death via the AKT signaling pathway [24, 25]. To explore the effect of glucose fluctuation on the AKT pathway in glomerular mesangial cells, we examined P-AKT, P-GSK-3β and cleaved caspase-3 expression levels in glomerular mesangial cells exposed to glucose fluctuation. As shown in Figure 4, there was no significant difference in the total AKT expression among the four groups. The level of P-AKT was lower in the HG group compared with that in the NG group, and the P-AKT level was lower in the FG group than that in the HG group. Moreover, high glucose could increase P-GSK-3β and caspase-3 activation in glomerular mesangial cells, but glucose fluctuation increase of P-GSK-3β and cleaved caspase-3 levels was more remarkable than stabilized glucose. These results indicated that glucose fluctuation accelerated glomerular mesangial cell apoptosis, which was linked to inhibiting decreased phosphorylation of AKT and increasing P-GSK-3β and cleaved caspase-3 levels.
Figure 4
Effects of glucose fluctuation on P-AKT, P-GSK-3b and cleaved caspase-3 levels. A – Representative bands of P-AKT and AKT. B – Representative bands of P-GSK-3b and GSK-3b. C – Representative bands of caspase-3. D – Analysis for optical density (OD) value of P-AKT, AKT, P-GSK-3b, GSK-3b and cleaved caspase-3 protein expression
The data are shown as mean ± SD and are expressed as fold difference vs. the NG group, bp < 0.05 vs. the HG group and cp < 0.05 vs. the FG group by one-way ANOVA.
Discussion
A previous in vitro study showed that glomerular mesangial cell apoptosis is a response to high glucose stimulation [26]. Furthermore, in an in vivo study, glomerular cell apoptosis could be found in the late phase of nephropathy [27]. In the present study, we chose a different time point to observe the mesangial cell vitality using the CCK8 kit, and we found that sugar fluctuation reduces mesangial cell vitality from 24 h, but has a significant reduction when compared to continuous high sugar stimulation from 48 h. Furthermore, we found that mannitol inhibited cell vitality at 72 h, so in the present study, we assessed the effect of glucose fluctuation on the mesangial cells and its related mechanism at 48 h. However, these results are different from a previous report [13]; we consider that this discrepancy may due to a different experimental protocol.
Overwhelming evidence indicated that high sugar induced cell apoptosis by increasing the generation of intracellular ROS in glomerular mesangial cells [5, 28]. Our previous study showed that fluctuating blood glucose increased intracellular ROS generation more than continuous high sugar; the SOD activity was obviously decreased and the MDA levels were increased in kidney subject to hyperglycemia in diabetic rats [23]. Furthermore, high sugar decreased the SOD activity and increased the MDA levels at 48 h compared with the normal glucose level, and blood glucose fluctuation markedly decreased their SOD activity and increased their MDA levels at 48 h. In addition, blood glucose fluctuation augmented apoptosis in pancreatic β cells through oxidative stress [20]. It is worth mentioning that our data suggested that glucose fluctuation accelerated cell apoptosis by an oxidative stress-dependent mechanism, and this result was consist with the present studies [29–32]. Hence, partly, we concluded that glucose fluctuation increased glomerular mesangial cell apoptosis via an oxidative stress mechanism.
The study confirmed that blood glucose fluctuation obviously decreased P-AKT action and increased the level of P-GSK-3β and Cleaved caspase-3 in glomerular mesangial cells compared to high sugar conditions. High glucose evokes an intrinsic pro-apoptotic signaling pathway in glomerular mesangial cells [6]. AKT activation was combined with protein, GSK-3β, which inhibits its kinase activity [33]. Studies have shown that P-GSK-3β was activated when there was inhibition of AKT phosphorylation in mesangial cells [34]. Especially, high sugar could decrease AKT activation and increased its downstream apoptosis related factors, such as GSK-3β and cleaved caspase-3, resulting in glomerular mesangial cell apoptosis [19]. In this study, glucose fluctuation could obviously decrease the P-AKT level and increased the expression of P-GSK-3β and cleaved caspase-3 in glomerular mesangial cells compared to high sugar conditions. On the basis of these results and previous work, mesangial cell apoptosis may be partly involved in inhibition of the AKT signaling pathway. These result demonstrate that the AKT signaling pathway may exert an important effect on glomerular mesangial cell apoptosis induced by unstable glucose stimulation.
In conclusion, blood glucose fluctuation increased mesangial cell apoptosis. This result may be partly induced by activating oxidative stress and inhibiting the AKT signaling pathway. These data provided important empirical evidence for us to understand the harm of blood glucose fluctuation.