Introduction
For millennia, plants have been an amazing pool of therapeutically important metabolites [1]. Although many currently used drugs are of plant origin, plants continue to serve as a source of more drugs or drug leads [2]. The use of plant-derived drugs dates back to the 19th century and since then many phytochemicals have been used in cancer treatment [3]. One of the benefits of using plant-derived drugs is their lower amount of side effects in comparison to their synthetic counterparts [4]. Based on structural affiliations, phytochemicals have been categorized into different groups. Out of these coumarins have shown great promise as anticancer agents and many plant-derived coumarins have been revealed to curb the proliferation of cancer cells via multiple mechanisms [5]. Bergamottin is a member of the furanocoumarin family of plant secondary metabolites, which has shown enormous potential as anticancer agents [6]. A number of studies have shown that bergamottin suppresses the proliferation and metastasis of cancer cells [7]. For instance, bergamottin blocks a wide array of oncogenic signalling pathways to suppress the proliferation of HepG2 cancer cells [8]. In another study bergamottin was reported to suppress the growth and invasion of glioma cells [9]. In yet another study, bergamottin was shown to promote apoptosis of human chronic myelogenous leukaemia [10]. Nonetheless, the growth inhibitory effects of bergamottin have not been thoroughly investigated in colon cancer. This study was therefore undertaken to investigate the anticancer effects of bergamottin against colon cancer together with elucidation of the underlying mechanisms. Ranked as the third most prevalent type of cancer, colon cancer imposes huge disease and economic burden on the world population. With 1.4 million cases of colon cancer diagnosed annually, colon cancer causes enormous human mortality and is hence considered as the fourth major reason for cancer-related mortality [11]. In 2013, around 0.7 million deaths were reported to be due to colon cancer worldwide [12]. Although the incidence of some cancers has declined to some extent it is believed the incidence of colon cancer will increase by 60% by 2030 [13]. The late diagnosis and dearth of potent and safe chemotherapeutic drugs are obstacles in the treatment of colon cancer [14].
Herein, we report that bergamottin suppresses growth of human colon cancer cells both in vivo and in vitro by induction of apoptosis and cell cycle arrest via deactivation of the Ras/Raf/ERK signalling pathway. Bergamottin may prove essential in the treatment of colon cancer and deserves further investigations.
Material and methods
Cell culture conditions
The colon cancer cell lines HT-29, SW-948, RKO and SW480, and normal CDD-18Co cells were obtained from the Cancer Research Institute of Beijing (Beijing, China) and maintained in Dulbecco’s modified Eagle’s medium (Invitrogen Life Technologies, Massachusetts, USA) supplemented with 10% fetal bovine serum (Invitrogen Life Technologies, Massachusetts, USA), 100 μg/ml streptomycin and 100 U/ml penicillin G (Himedia, Pennsylvania, USA) in an incubator at 37°C with 5% CO2.
Cell proliferation assay
The viability of the human colon cancer cells and normal astrocytes was monitored by WST-1 assay. In brief, HT-29 and RKO cells were cultured in 96-well plates at the density of 2 × 105 cells/well and treated with 0 to 200 μM concentrations of bergamottin for 24 h at 37°C. This was followed by the incubation of the cells with WST-1 at 37°C for 4 h. The absorbance was then measured at 450 nm using a Victor3 microplate reader to determine the proliferation.
Apoptosis assay
The HT-29 and RKO cells (0.6 × 106) were cultured in 6-well plates and treated with bergamottin at the concentrations of 0, 3.12, 6.2 and 12.5 μM for 24 at 37°C. The cells were then subjected to phosphate buffered saline (PBS) washing and subsequently stained with DAPI. Finally, the cells were examined under a microscope to detect the induction of apoptosis. For determination of the percentage of apoptosis the transfected cells were stained with annexin V/PI and subsequently examined by a flow cytometer.
Cell cycle analysis
The cultured human colon cancer HT-29 and RKO cells were firstly treated with varied concentrations of bergamottin n for 24 at 37°C. The cells were then washed with PBS. Afterwards, the HT-29 and RKO cells were stained with propidium iodide (PI) and the distributed of the cells in cell cycle phases was assessed by a FACS flow cytometer.
Wound-healing assay
The bergamottin treated colon cancer cells were cultured at 37°C until confluence (80%). This was followed by removal of the media and subsequent washing of the cells with PBS. A scratch was made with a pipette tip and a photograph was taken. The plates were then kept at 37°C for 24 h and a photograph was taken again under a microscope.
Cell invasion assay
The effects of bergamottin on the invasion ability of HT-29 and RKO cells was determined by transwell chambers with Matrigel. Around 200 ml cell cultures were placed onto the upper chambers and only medium was placed in the bottom wells. After 24 h of incubation, the cells were removed from the upper chamber and the cells that invaded via the chambers were subjected to fixation with methyl alcohol and subsequently stained with crystal violet. An inverted microscope was used to count the number of invaded cells at 200X magnification.
Western blot analysis
The HT-29 and RKO cells were cultured for 24 h at 37°C. Next the cultures were transferred to 50 ml centrifuge tubes and centrifuged at 12 000 rpm for 10 min. The pellets were washed with PBS and then resuspended in lysis buffer (20 mM HEPES, 350 mM NaCl, 20% glycerol, 1% Nonidet P 40, 1 mM MgCl2, 0.5 mM EDTA, 0.1 mM EGTA, 1 mM DTT, 1 mM PMSF, protease inhibitor cocktail, and phosphatase inhibitor cocktail). BCA assay was used to determine the concentration of the proteins in each sample. Equal concentrations of the proteins from each sample were loaded and separated on the SDS-PAGE and then transferred to polyvinylidene fluoride membrane. These membranes were then treated with TBS and then exposed to primary antibody (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at 4○C for 24 h. Afterwards, the cells were treated with HRP-conjugated anti-rabbit secondary antibody and subsequently chemiluminescence reagent was used for the visualisation of the proteins.
In vivo study
National Institutes of Health standards for the care and use of laboratory animals and approved by Cancer Hospital of China Medical University under approval number CMU67/34/2018. The mice (4 weeks old, weighing around 24 ±2.2 g) were injected with 5 × 106 HT-29 cells subcutaneously at the left flank. As the tumours were apparent, the mice (n = 5) for each group were injected intraperitoneally with DMSO (0.1%) dissolved bergamottin and diluted with 100 μl of normal saline at 0, 15, 30 and 60 mg/kg body weight and taken as the first day of the experiment. Bergamottin was given to the rats thrice a week while the control mice were given DMSO (0.1%) in normal saline only. At the end of 6 weeks, the mice were sacrificed and tumours were harvested for assessment of tumour growth and other investigations.
Results
Bergamottin inhibits the proliferation of human colon cancer cells
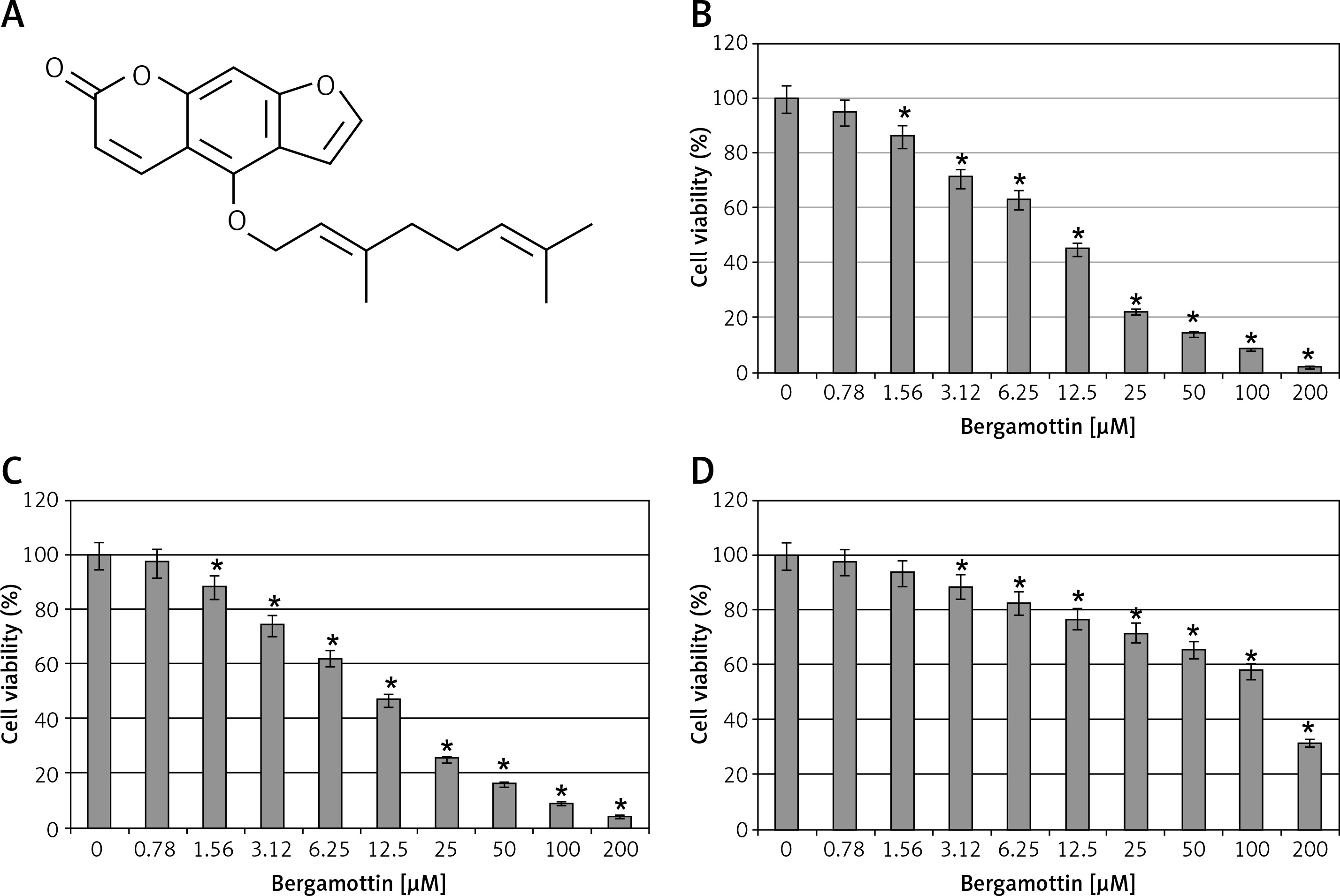
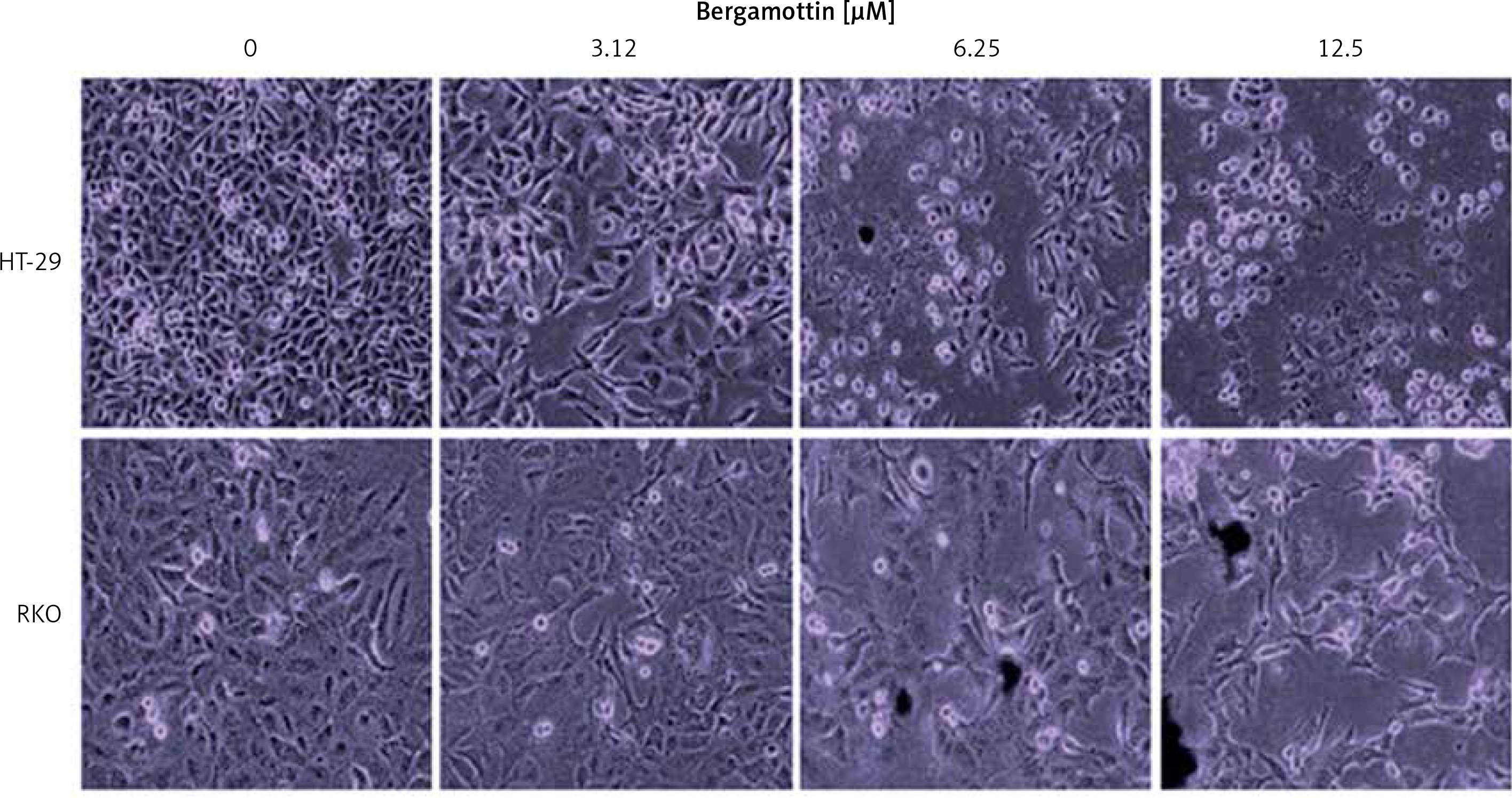
The antiproliferative effects of bergamottin were evaluated by MTT assay on different human colon cancer cell lines. Bergamottin treatment resulted in a significant decrease in the viability of all the colon cancer cell lines and the IC50 varied from 12.5 to 30 μM (Table I). Nonetheless, more profound growth inhibitory effects were observed against the HT-29 and RKO cells as evident from the IC50 of 12.5 μM for bergamottin against these two cell lines (Figure 1 B). Next, the cytotoxic effects of bergamottin were also investigated on the normal CDD-18Co cells and interestingly the toxic effects were comparatively much lower on the normal cells as evident from the IC50 of > 100 μM (Figure 1 B). Microscopic analysis showed that bergamottin induced significant changes in the morphology of both HT-29 and RKO cells such as shrinkage, membrane blebbing and rounding (Figure 2).
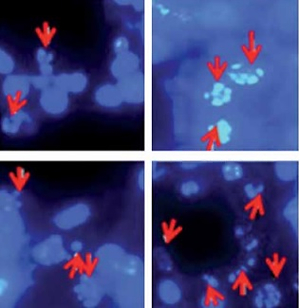
Bergamottin triggers apoptosis in RKO and HT-29 cells
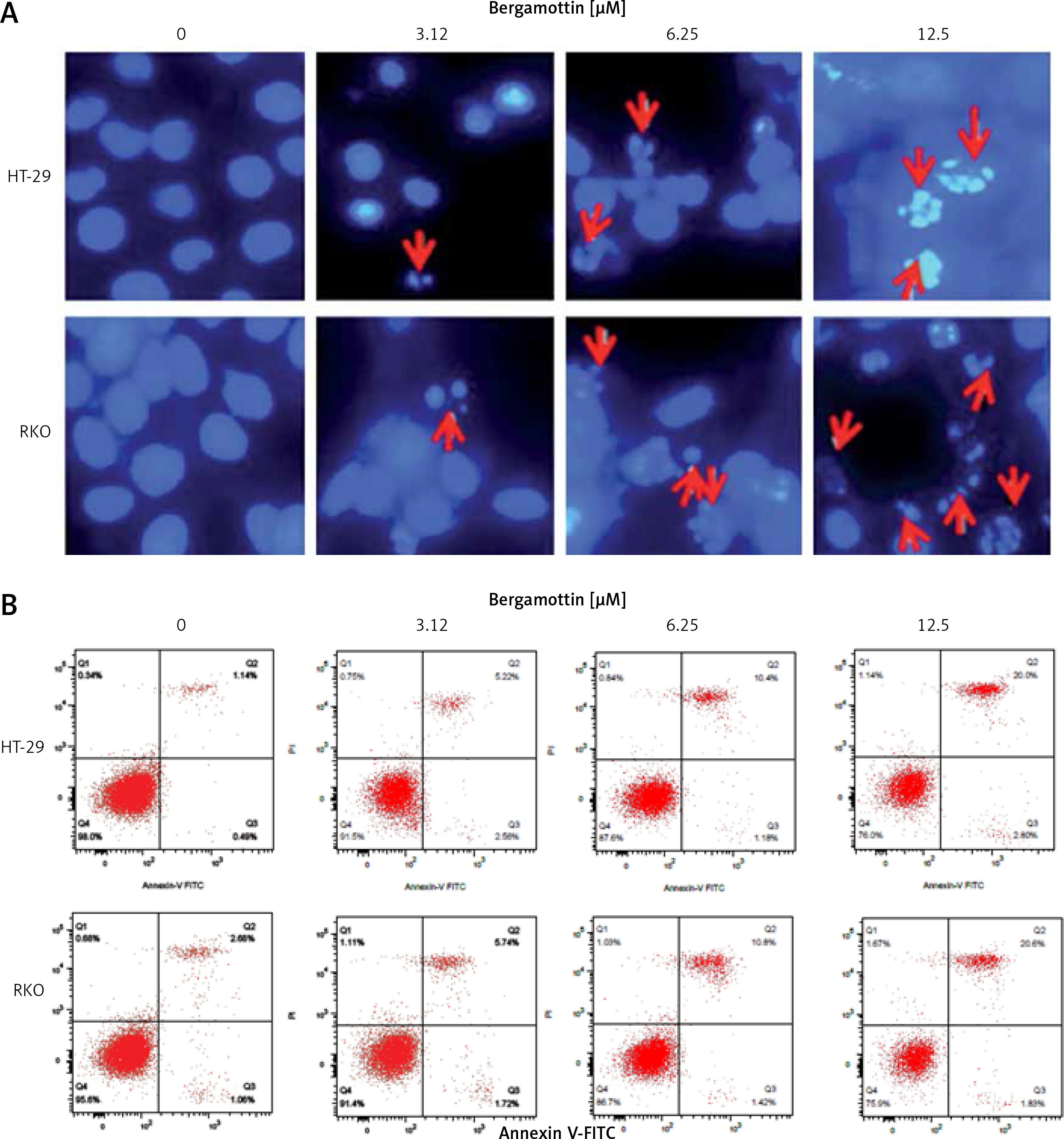
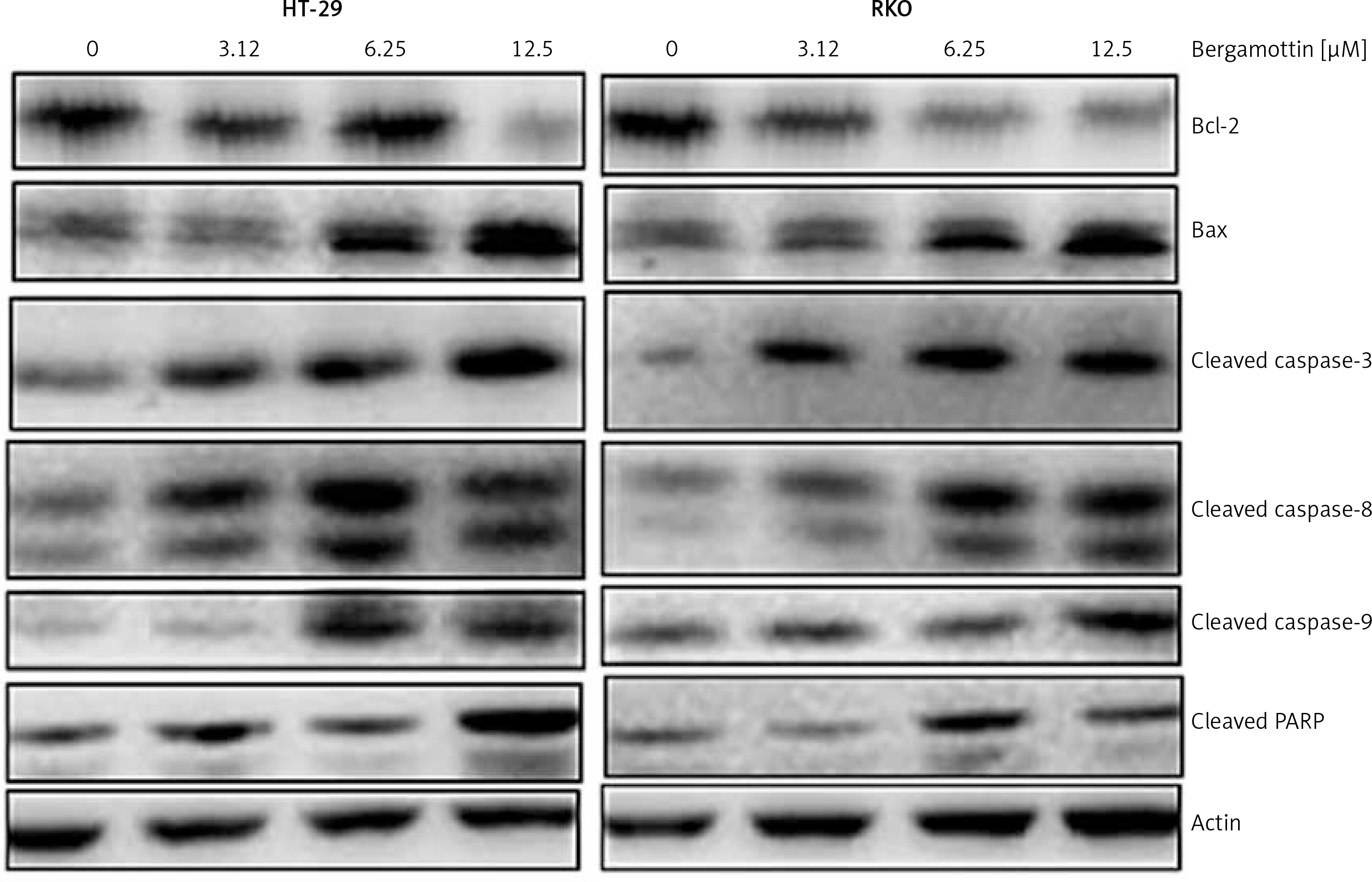
To reveal the reasons behind the anticancer effects of bergamottin, DAPI and annexin V/PI staining of the bergamottin treated HT-29 and RKO cells were performed. DAPI staining showed clear nuclear fragmentation indicative of apoptosis (Figure 3 A). Annexin V/PI staining revealed that the apoptotic HT-29 and RKO cell percentage increased upon bergamottin treatment. The percentage of HT-29 apoptotic cells was 2.05, 7.20, 12.62 and 28.9% and that of RKO apoptotic cells was 2.99, 6.5513.79 and 26.01% at 0, 3.12, 6.25, 12.5 and 25 μM concentration of bergamottin (Figure 3 B). Western blotting was further used to confirm the apoptosis and the results showed that bergamottin significantly increased Bax expression, which was concomitant with depletion of Bcl-2 expression (Figure 4). Moreover, bergamottin also caused an increase in the cleavage of caspase-3, 8 and 9 in a concentration-dependent manner. These results suggest that bergamottin suppresses the growth of colon cancer cells at least partially via promotion of apoptosis.
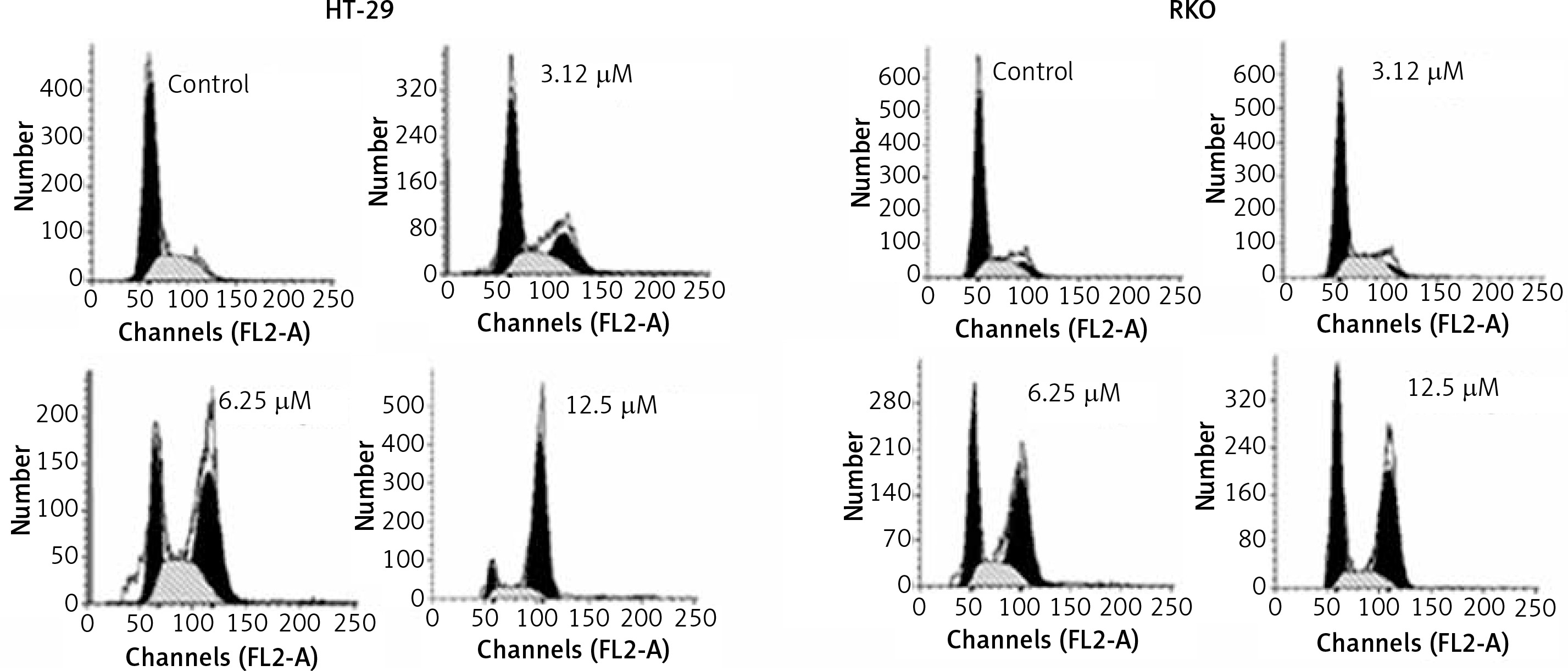
Bergamottin induces G2/M cell cycle arrest of HT-29 and RKO cells
Cell cycle analysis of the bergamottin treated HT-29 and RKO cells was also performed to assess whether bergamottin triggers cell cycle arrest. The results showed that bergamottin caused an increase in the percentage of the G2/M phase cancer cells. The HT-29 and RKO increased from 1.21% and 0.75% in control to 87% and 61% at 12.5 μM concentration (Figure 5). Next, western blot analysis was performed to determine the expression of the regulatory proteins of cell cycle and it was revealed that bergamottin caused significant reduction in Cdc2, cyclin A, and cyclin B. Nonetheless, the levels of negative regulatory proteins of cell cycle progression (p21Cip1, and p27Kip1) were remarkably and dose-dependently depleted by bergamottin (Figure 5).
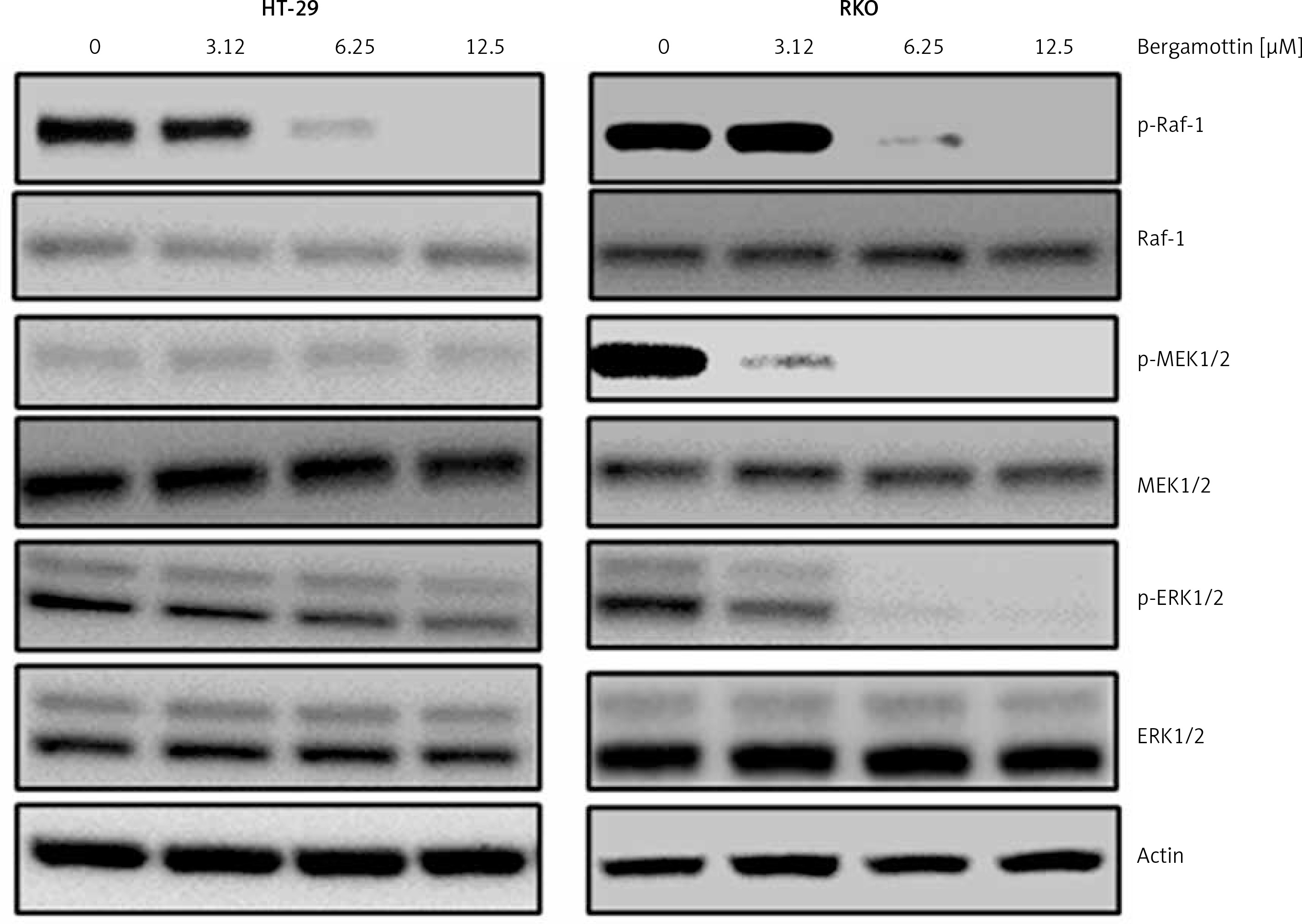
Bergamottin blocks Raf/MEK/ERK signal transduction
The effect of bergamottin was also assessed on the Raf/MEK/ERK signalling cascade to ascertain whether bergamottin induced G2/M arrest is via blocking of this pathway. It was found that bergamottin prompted concentration dependently suppression of the phosphorylation of Raf-1, MEK1/2 and ERK1/2 with no visible impact on Raf-1, MEK1/2 and ERK1/2 expression (Figure 6).

Bergamottin inhibits invasion of HT-29 and RKO cells
Transwell assays were performed to determine the effects of bergamottin on the invasion of the HT-29 and RKO cells. The results showed that bergamottin could decrease the invasion of the RKO and HT-29 colon cancer cells considerably (Figure 7 A). Additionally, the decrease of cell invasiveness was also associated with depletion of metalloproteinase 2 and 9 expression (Figure 7 B).
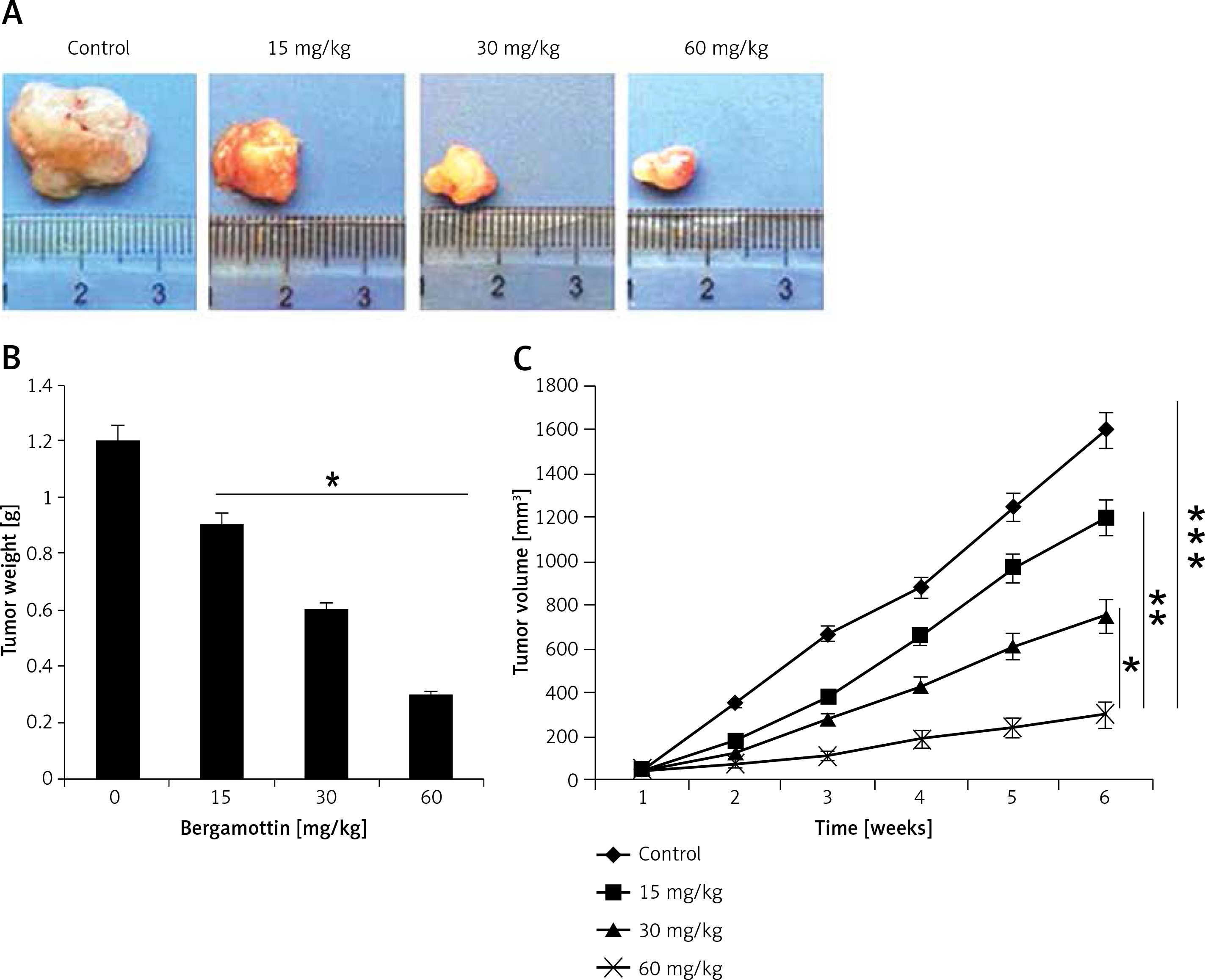
Bergamottin suppresses xenografted tumour growth in vivo
To assess the in vivo anticancer effects of bergamottin, a xenografted mouse model was used. The results showed that bergamottin caused reduction in the xenografted tumour growth (Figure 8 A). The tumour weight was significantly decreased and the tumour volume showed a constant reduction over the period of 6 weeks (Figure 8 B). Additionally, no apparent toxicity was observed on the mice during the in vivo study (Figure 9).

Discussion
Colon cancer is a devastating cancer and its incidence is expected to escalate significantly in the next few decades [15]. The unsatisfactory clinical outcome, flawed treatment strategies and adverse effects of the existing drugs form the major hurdles in colon cancer treatment. Moreover, emergence of chemoresistance and frequent relapse make it more difficult to treat colon cancer [11]. Herein, the anticancer effects of a naturally occurring furanocoumarin, bergamottin, were evaluated against a panel of human colon cancer cell lines. Although bergamottin inhibited the growth of all the colon cancer cell lines, the IC50 values were found to be lowest against the HT-29 and RKO cells. Additionally, the cytotoxicity of bergamottin was found to be very low against the normal CDD-18Co cells. These findings are consistent with previous studies wherein bergamottin was found to suppress the growth of cancer cells by blocking the STAT3 signalling pathway [16]. Similarly, another related coumarin, auraptene, has been shown to curb the growth of gastric cancer cells via induction of apoptosis [17]. Apoptosis is a systematic and orchestrated process in living cells that enables the elimination of harmful or defective cells from the body. Apoptosis has gained tremendous attention for its therapeutic implications in cancer [18]. Many anticancer agents halt the growth of cancer cells by promoting apoptosis [19]. In this study, we found that bergamottin induces apoptotic cell death of the HT-29 and RKO colon cancer cells. The expression of the important apoptotic biomarker proteins Bax and Bcl-2 [20] was also altered upon bergamottin treatment. Additionally, bergamottin enhances the cleavage of caspase-3, 8 and 9 as well as that of PARP, favouring apoptosis. In addition to apoptosis several plant-derived molecules have also been shown to inhibit cell division by arresting the cells at different checkpoints [21]. Herein we also observed that bergamottin caused the arrest of the HT-29 and RKO cells at the G2/M phase of the cell cycle. This was also associated with depletion of Cdc2, cyclin A, cyclin B1 and upsurge of the negative regulators of cell cycle progression (p21Cip1, and p27Kip1). Studies have shown that inhibitors of the Raf/MEK/ERK signalling pathway may provide effective anticancer agents for the management of diverse cancer types [22, 23]. However, the currently used regimen of drugs is disappointingly ineffective to block this pathway. Therefore the effects of bergamottin on this pathway were examined and it was found that bergamottin could block this pathway. Bergamottin also suppressed the migration and invasion of the colon cancer cells, which is agreement with previous studies wherein bergamottin has been reported to inhibit the invasion of glioma cells [9]. Given the promising results of the in vitro anticancer effects of bergamottin, xenografted tumour mouse models were used to evaluate the anticancer effects of bergamottin in vivo. The results showed that bergamottin could inhibit the tumour growth in vivo, indicative of the anticancer potential of bergamottin.
In conclusion, the findings of the present study indicate that bergamottin exerts anticancer effects on human colon cancer cells via induction of apoptosis and cell cycle arrest. Bergamottin could also suppress invasion of colon cancer cells via blocking the Raf/MEK/ERK signalling cascade. All considered, bergamottin may prove beneficial in colon cancer treatment and warrants further research endeavours.